




Engineered bacterial extracellular vesicles as next-generation precision postbiotics: strategies, challenges and prospects
 0
0 Abstract
Postbiotics, defined as inactivated microorganisms and/or their components conferring a health benefit, represent a paradigm shift from traditional probiotics by offering superior safety, stability, and precise dosing. Bacterial extracellular vesicles (BEVs), natural nanoparticles secreted by bacteria, are key mediators of microbiota-host communication. BEVs have emerged as promising natural postbiotic agents due to their ability to transport bioactive cargo and modulate host physiology. However, the therapeutic application of natural BEVs (nBEVs) is constrained by limitations, including potential safety concerns, lack of target specificity, low yield, and compositional heterogeneity. To overcome these barriers, the concept of engineered BEVs (eBEVs) has been introduced. By employing genetic modification of parent bacteria, surface functionalization, and tailored cargo loading, eBEVs can be precisely designed. This engineering transforms nBEVs into advanced, targeted delivery platforms that significantly expand the scope and efficacy of postbiotics. This review explores the vast potential of eBEVs as next-generation precision postbiotics to treat multiple disorders. Despite the transformative potential, translating eBEVs into clinical practice faces challenges. These include scalable manufacturing processes, comprehensive in vivo safety and pharmacokinetic profiling, and the establishment of regulatory frameworks. Future progress hinges on interdisciplinary efforts to develop smart, stimulus-responsive BEVs, enhance endosomal escape efficiency, and conduct rigorous clinical trials. Ultimately, eBEVs stand at the forefront of a new era in microbiota-based therapeutics, merging the biocompatibility of natural vesicles with the precision of synthetic nanomedicine.
Keywords
INTRODUCTION
The microbiome refers to the collection of bacteria, fungi, and other microorganisms, identified through metagenomics and meta-transcriptomic analysis, that live in the gut, other human mucosae (e.g., oral, nasal, vaginal mucosae), and on the skin surface. Breakthrough research on the microbiome is opening new avenues for disease treatment[1]. While traditional probiotics can exert health benefits by positively modulating the immune system, competitively inhibiting pathogens, enhancing intestinal barrier function, and regulating lipid metabolism[2], their application is significantly limited by the difficulty that microbes face in maintaining viability and colonize the gastrointestinal tract (GI tract), and inconsistent effects[3]. As a consequence, the concept of postbiotics has emerged in the microbiome field. According to the definition of the International Scientific Association for Probiotics and Prebiotics (ISAPP), postbiotics are preparations of non-living microorganisms and/or their components that are beneficial to host health[4], which can address the limitations of probiotics. Studies have shown that postbiotics can even be more effective than live bacteria in certain cases, such as inactivated Akkermansia muciniphila (A. muciniphila), which demonstrates superior performance in increasing GLP-1 levels and improving liver function[5]. This transformation marks the shift from relying on the activity of microorganisms for broad-spectrum regulation to an era of “non-living preparations” characterized by precision, stability, and safety.
Extracellular vesicles (EVs) are non-replicating, lipid bilayer-enclosed nanoparticles released by cells, with diameters ranging from 20-400 nm. They carry various biologically active components, including DNA, RNA, lipids, proteins, and metabolites[6]. EVs released by bacteria are commonly referred to as bacterial-derived extracellular vesicles (BEVs)[7]. They represent one of the key underlying mechanisms behind the detrimental or beneficial effects of many pathogenic, commensal, and probiotic bacteria[8]. Among them, those derived from Gram-negative (G-) bacteria are called outer membrane vesicles (OMVs), while those derived from Gram-positive (G+) bacteria are defined as cytoplasmic membrane vesicles (CMVs)[9].
Current research shows that BEVs represent novel potential postbiotic candidates with health benefits[10]. BEVs are capable of penetrating intra- and extra-cellular barriers, mediating extensive crosstalk with mammalian cells, and delivering their cargo to the host[8,11]. BEVs possess inherent postbiotic properties. They can mediate the interaction between the microbiome and the host, which has attracted widespread attention and is under intensive investigation[12]. These are referred to as natural BEVs (nBEVs).
However, the application of nBEVs as postbiotics carries certain risks, as they may contain uncontrolled components like toxins and immunogenic substances, potentially leading to safety concerns[13]. BEVs derived from G- bacteria consistently contain lipopolysaccharide (LPS), a pro-inflammatory activator, triggering severe host immune responses[14-16]. Some BEVs are associated with pathogenesis and serve as long-distance delivery vehicles for virulence factors related to adhesion, invasion, antimicrobial resistance, and host immune regulation[17]. Additionally, due to the inability to fully control and standardize the cultivation conditions, batch-to-match differences of nBEVs lead to heterogeneity[18]. The poor targeting ability and low yield of nBEVs also hamper the clinical application[19,20]. These uncontrolled components and variability present potential safety challenges for the postbiotic application of nBEVs.
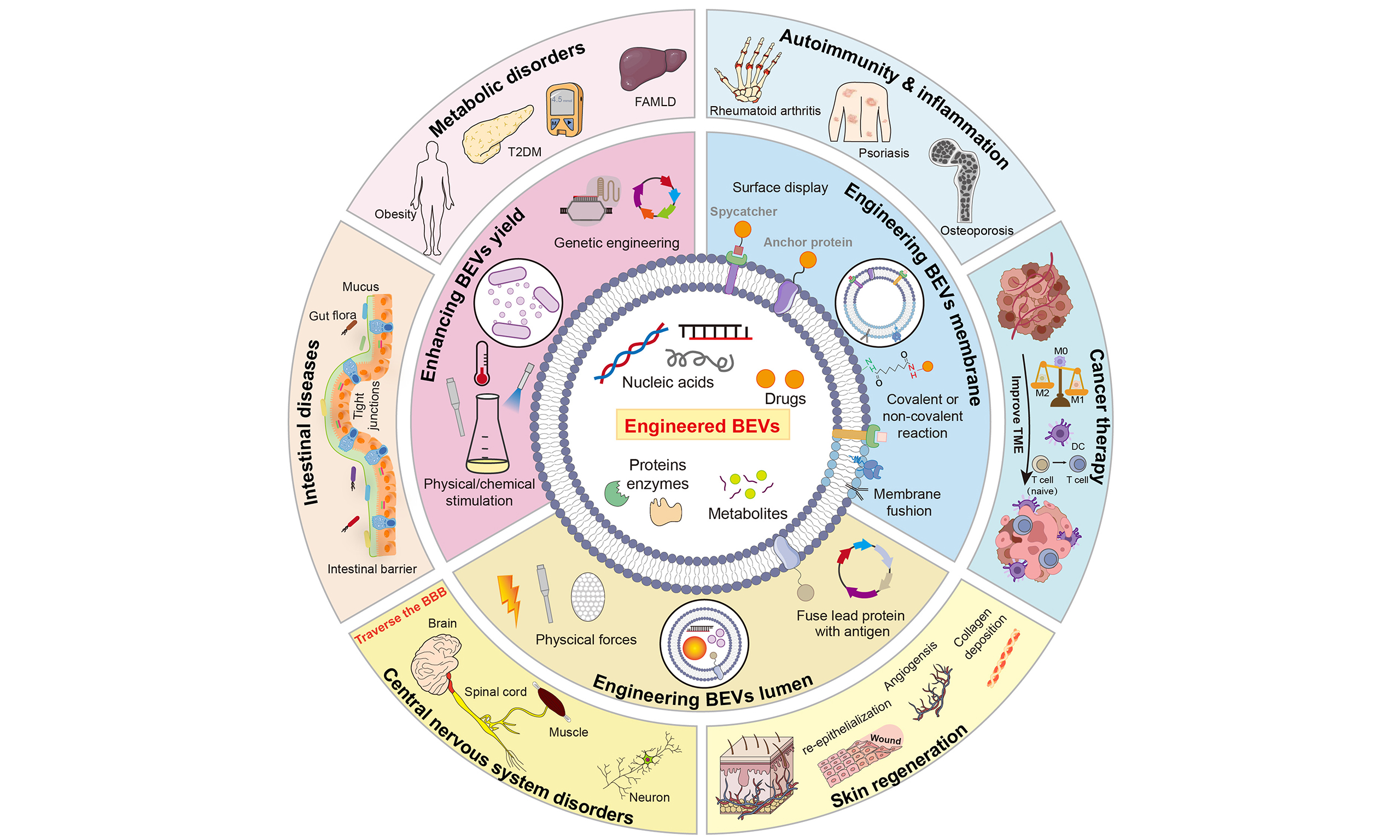
In order to better utilize BEVs, enhancing security, improving efficacy and new functions, and achieving precise delivery, diverse engineering approaches can be employed to modify nBEVs. Main approaches include (i) modifying toxin genes; (ii) controlling the expression of exogenous proteins; (iii) surface modification of proteins or peptides for targeting purposes; (iv) physical methods such as membrane fusion, ultrasound, and squeezing wrapping, combining with nanoparticles; and (v) regulating genes involved in BEV synthesis to enhance BEV yields[21-23]. Unlike traditional probiotics and postbiotics, engineered BEVs (eBEVs) are engineered to achieve four distinct levels of precision: (i) specific targeted delivery via surface engineering; (ii) controlled and stimuli-responsive cargo release; (iii) programmable therapeutic payloads tailored to specific diseases; (iv) reduction of off-target toxicity through detoxification. Also, the therapeutic effects of traditional probiotics and postbiotics are difficult to fully predict or standardize because of the complex and undefined components of bacteria. In contrast, eBEVs are welldefined nanoscale vesicles that emerge as postbiotics of the next generation, which can be precisely engineered at the genetic, physical, and chemical levels, while retaining natural biocompatibility and intrinsic bioactivity. Collectively, eBEVs, as next-generation precision postbiotics, offer a level of control that cannot be achieved by unmodified nBEVs or other probiotics/postbiotic formulations [Figure 1]. This review will systematically elaborate on the strategies to design and construct eBEVs, their application prospects in various diseases, and the challenges in their clinical translation.
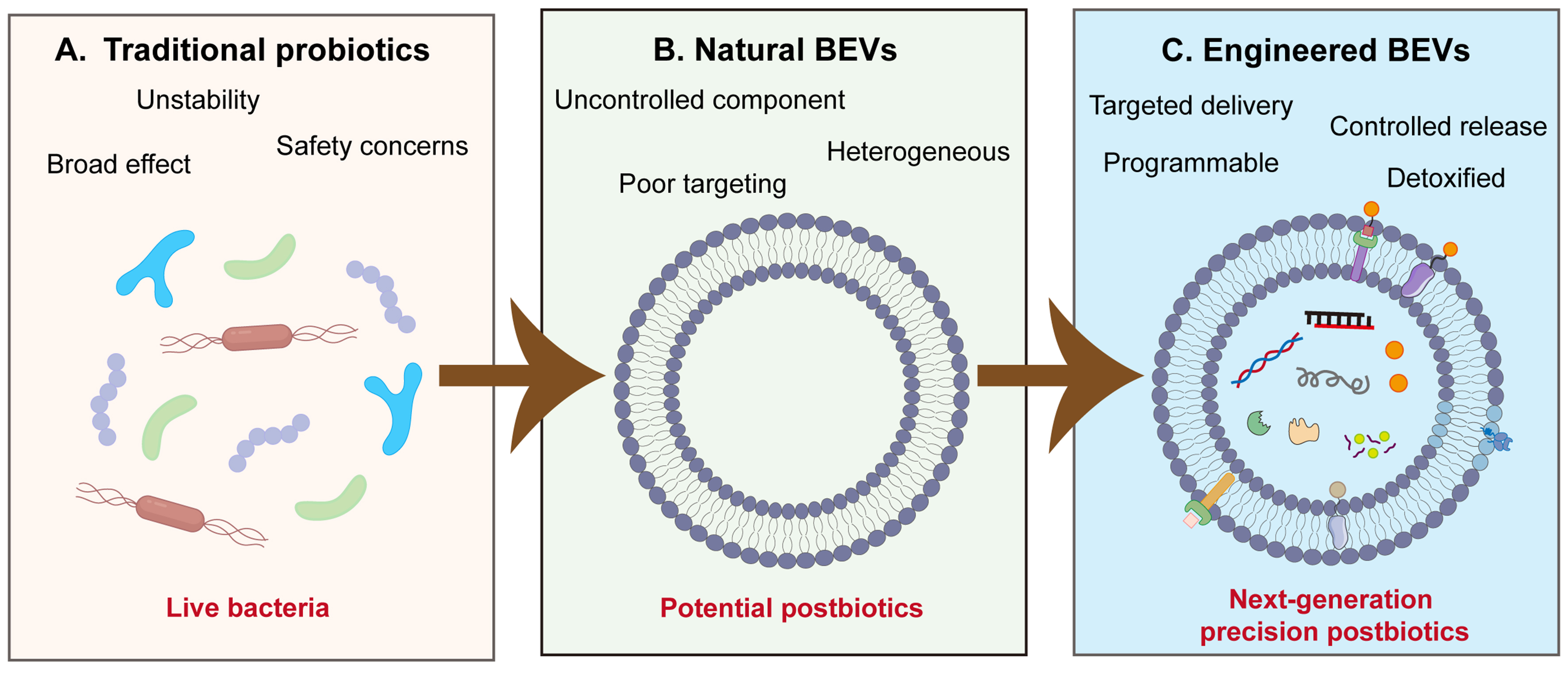
Figure 1. Paradigm shift from traditional probiotics to eBEVs as next-generation precision postbiotics. (A) Traditional probiotics: live beneficial bacteria but with limited stability, potential safety risks, and non-specific broad-spectrum effects; (B) nBEVs: improve general host health but have remaining issues with composition, safety, and stability; (C) eBEVs: precise platform with targeted delivery, controlled release, programmable payload loading, and detoxified profiles. BEVs: Bacterial extracellular vesicles; nBEVs: natural bacterial extracellular vesicles; eBEVs: engineered bacterial extracellular vesicles.
OVERVIEW OF nBEVs
G+ bacteria have thick cell walls rich in peptidoglycan (PG) and teichoic/lipoteichoic acids (TA/LTA), whereas G- bacteria have inner and outer membrane structures rich in LPS, lack of TA/LTA, and a lower content of PG. Both types of bacteria can spontaneously form and secrete BEVs[24]; however, the formation mechanisms differ due to the different cell structures. G- bacteria mainly secrete BEVs through two pathways: the one pathway produces OMVs that contain only the components of the outer membrane. The other pathway, resulting from cell wall degradation leads to explosive outer membrane vesicles (EOMVs) or outer-inner membrane vesicles (OIMVs) that contain inner and outer membranes as well as cytoplasmic components of the[9,16]. G+ bacteria, due to their thicker PG layer, were previously believed to be unable to secrete BEVs. However, not until 2009 were G+ bacteria discovered to produce vesicles through a process similar to “bubble-like cell death”[25]. The specific mechanism is still unclear and may be related to the action of autolysins or phagocytic lysis enzymes[26].
Oral administration of BEVs and their absorption through the intestines is the most prevalent route. BEVs carry numerous miRNAs, proteins, and lipids from parental cells, enabling them to traverse the intestinal barrier, transfer bioactive molecules into intestinal epithelial cells, and effectively diffuse into the circulatory system for transmission to peripheral organ systems[27,28]. BEVs administrated via surface routes require the ability to penetrate bodily barriers and reach host cells. Within 24 h, BEVs from Lactobacillus paracasei can reach the deepest part of the dermis and alleviate tumor necrosis factor-α (TNF-α)-induced skin inflammation[29]. Moreover, BEVs can enter respiratory epithelial cells and release their contents to stimulate lung immune cells while inducing protective immune responses[30].
BEVs possess a wide range of functions, contingent upon their unique nanostructure and the diverse biomolecules they carry. To date, BEVs have been confirmed to exert broad health-promoting effects [Figure 2 and Table 1]. A large body of research has explored the beneficial effects of BEVs in local intestinal environment, including enhancing the intestinal barrier integrity[45], modulating the gut microbiota[46,47], and regulating local inflammatory responses[48]. In addition, BEVs exhibited systemic health effects, such as improving brain health[49,50], regulating anti-tumor immune responses[51,52], and alleviating metabolic diseases[33,34]. These studies indicate that nBEVs are potentially vital regulators of general health and may provide protective effects against diseases. However, issues such as safety, stability, and scalability require further refinement.
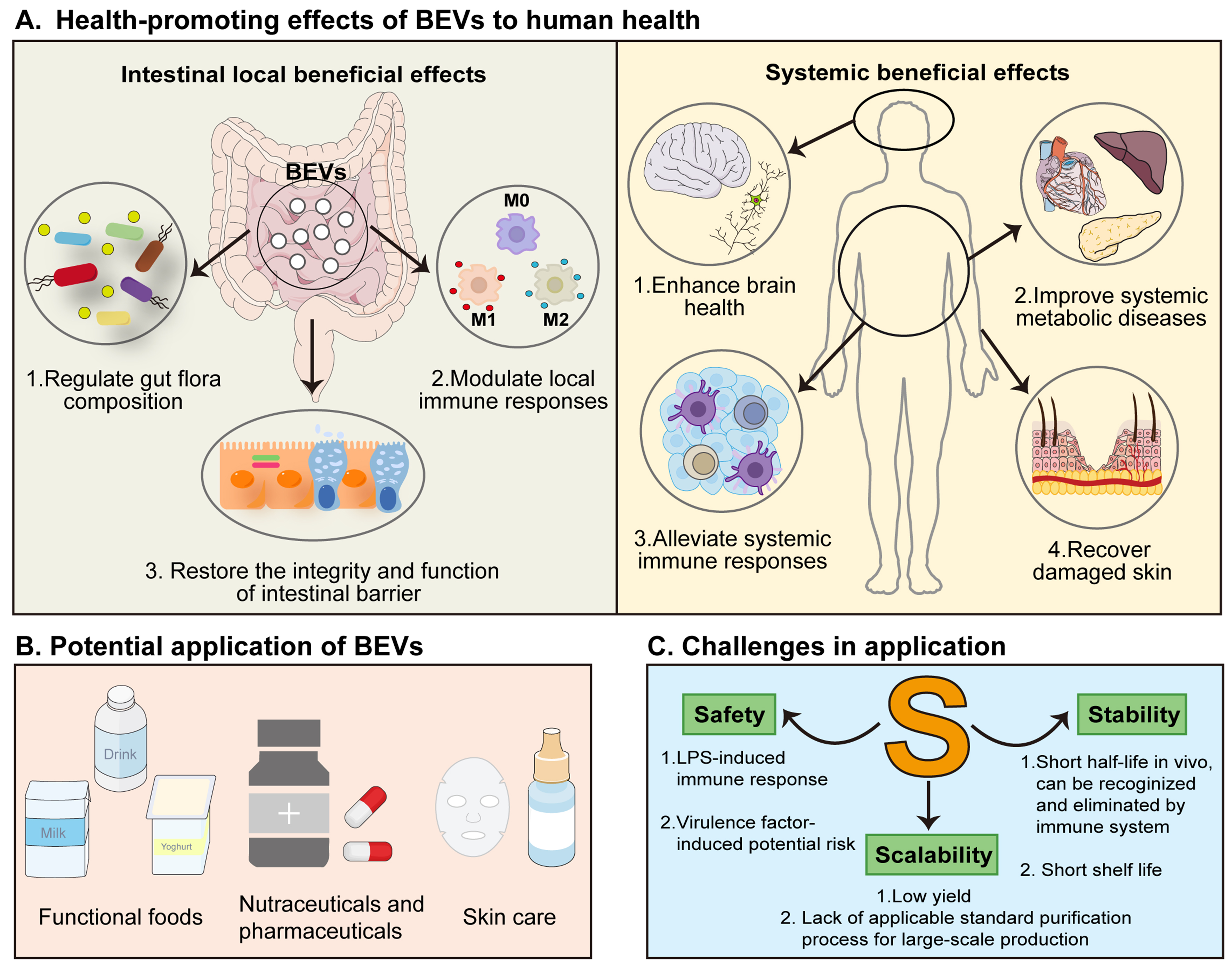
Figure 2. Health-promoting effects of nBEVs and potential challenges in their applications. (A) BEVs exert their health-promoting effects on the intestinal tract by regulating the composition of the host’s intestinal microbiota, modulating local immune responses in the intestine, and restoring the integrity and function of the intestinal barrier. Furthermore, BEVs also have a positive impact on the overall health of the host (including the brain, metabolic diseases, immune system, and skin); (B) Potential applications of BEVs as postbiotics in the food and biomedical industries; (C) Current challenges for the safe and effective clinical application of BEVs. BEVs: Bacterial extracellular vesicles; nBEVs: natural bacterial extracellular vesicles; LPS: lipopolysaccharide.
nBEVs derived from different bacterium and their therapeutic effects
| Bacterium | Model | Mechanism | Effects | Ref. |
| Lysobacter enzymogenes C3 | F. subglutinans Infection | Deliver antifungal compounds to treat fungal diseases | [31] | |
| Bacteroides longum KACC 91563 | Diarrhoea due to food allergy | Induce apoptosis of BMMCs,prevention of food allergy | [32] | |
| A.muciniphila MucT ATCC BAA-835 | HFD induced diabetic mice | Increased tight junction protein expression in Caco-2 cells and ultimately improved intestinal barrier integrity | [33] | |
| A. MucT ATCC BAA-835 | HFD induced fat mice | Improved the intestinal barrier integrity, inflammation, energy balance, and blood lipid profile and glucose level | [34] | |
| Escherichia coli Nissle 1917 | DSS induced mouse colitis | ↓ Body weight loss ↓ Disease activity index | Reduced DSS-induced weight loss and ameliorated clinical symptoms and histological scores | [35] |
| Lactobacillus animalis ATCC 35046 | GC-induced ONFH | Enhancing blood vessel abundance, maintaining osteogenic activity, reducing cell apoptosis, and attenuating trabecular bone and marrow damage, thus protecting the mice from GC-induced ONFH | [36] | |
| A.muciniphila | DSS-induced colitis | ↓ IL-6 | Protected DSS-induced IBD phenotypes, such as body weight loss, colon length, and inflammatory cell infiltration of colon wall | [37] |
| Bacteroides fragilis | TNBS-induced experimental colitis | Induce immunomodulatory effects and prevent experimental colitis | [38] | |
| Clostridium butyricum | DSS-induced colitis in mice | Inhibition of NF-kB pathway activation Improving the intestinal barrier | Alleviate bacterial dysbiosis,improve intestinal barrier integrity and inhibit the inflammatory response | [39] |
| A.muciniphila | HFD/CCL4-induced liver injury | Improvement in gut health Anti-inflammatory effects on liver and adipose tissues | [40] | |
| LGG | ALD | Mediated by intestinal AhR signaling ↑ Intestinal interleukin‐22‐Reg3 ↑ Nrf2-tight junction signaling pathways | Protected the intestine from alcohol‐induced barrier dysfunction and the liver from steatosis and injury | [41] |
| Lactobacillus paracasei | TNF-α-induced inflammatory phenotypes in human skin | Restored the TNF-α-induced epidermal malformation, abnormal proliferation of keratinocytes in the basal layer, and reduction in dermal collagen synthesis | [29] | |
| Lactobacillus plantarum | Skin aging | Modulated the mRNA expression of ECM related genes | Reduction in skin moisture content and increase skin density Suppressed wrinkle formation and pigmentation | [42] |
| Lactobacillus plantarum ZS62 | Alcohol-induced subacute liver injury | Reducing inflammation and enhancing antioxidant capacity | Attenuated alcohol-induced weight loss; Prevented morphological changes in hepatocytes; Reduced AST, ALT, Haase, PC III and inflammatory cytokines; Down-regulated inflammation-related genes and up-regulated lipid- and oxidative-metabolism genes | [43] |
| Lactobacillus plantarum DP189 | MPTP-induced PD mice | Activated the expression of Nrf2/ARE and PGC-1α pathways and suppressed the NLRP3 inflammasome | Increased the levels of SOD, GSH-Px, IL-10, decreased the levels of MDA, ROS, TNF-α, IL-6 and IL-1β. Reduced the α-SYN accumulation in SN. Reshaped the gut microbiota in PD mice | [44] |
ENGINEERING STRATEGIES FOR BEVs
To meet the clinical application requirements, multiple engineering strategies have been developed to modify nBEVs with enhanced yield, therapeutic efficacy, stability and targeting capacity. The modification methods for nBEVs engineering can be mainly classified into three core aspects, focusing on increasing the production of BEVs, modifying the membrane surface, and remolding of inner lumen[18]. This review thoroughly discusses the advanced approaches for BEVs engineering to address the current application bottlenecks.
Enhancing BEVs yield
Current research mainly focuses on enhancing the production of BEVs by genetically engineering of the parent strains, optimizing cultivation conditions and downstream processes [Figure 3]. Utilizing the various strategies presented below to increase BEVs yield could pave the way for their application.
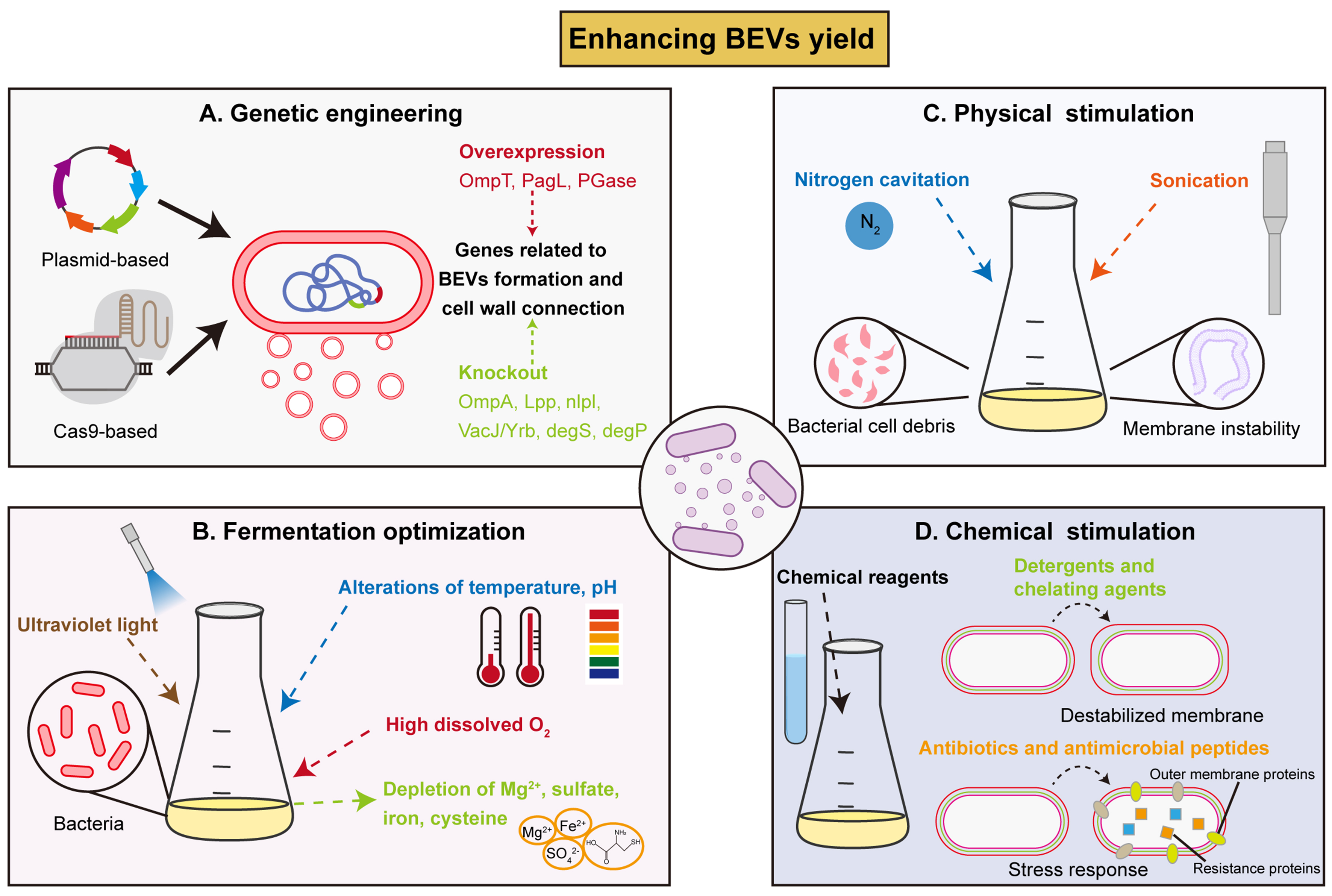
Figure 3. Strategies to enhance BEVs yield. (A) Utilization of genetic engineering techniques (recombinant plasmids, CRISPR-Cas9) to overexpress or knockout genes related to BEVs formation and cell wall composition; (B) Fermentation optimization, such as UV light stimulation, temperature/ pH alteration, and depletion of nutrients to enhance BEVs yield; (C) Physical stimulation with extreme environmental forces (nitrogen cavitation, sonication) to produce cell debris and induce membrane instability, thus enhance BEVs production; (D) Chemical reagents treatment (detergents, chelating agents, antibiotics) to induce more BEVs production as a self-protective response. BEVs: Bacterial extracellular vesicles; OmpT: outer membrane protease T; PagL: lipid A 3-O-deacylase PagL; PGase: peptidoglycan hydrolase; OmpA: outer membrane protein A; Lpp: murein lipoprotein; NlpI: lipoprotein NlpI; DegP: periplasmic serine endoprotease DegP; CRISPR-Cas9: clustered regularly interspaced short palindromic repeats-associated protein 9; UV: ultraviolet.
Genetic modification
Genetic engineering modification utilizes gene editing technology to regulate the expression of genes related to the biogenesis of BEVs to achieve large-scale generation of BEVs[53]. The formation of BEVs in G- bacteria (in which most studies have been performed) requires the detachment of the outer membrane from the PG layer and inner membrane, thus breaking the connections between the layers of the cell wall (i.e., inner membrane-PG-outer membrane structures) facilitates the release of BEVs[54].
The gene overexpression strategy is explored to enhance BEVs production. The outer membrane protease OmpT of enterohemorrhagic Escherichia coli (E. coli) (EHEC) influences the bacterium’s ability to adapt to the host environment by dynamically regulating the production, composition, and size of OMVs. Overexpression of the ompT significantly increases OMVs production with a 40-fold increase compared with the wild type and further modulates bacterial growth[55]. Another study reveals that lipid A (a component of the LPS) deacylation is a vital mechanism driving OMVs biogenesis. Overexpression of deacylase PagL in Salmonella typhimurium induces membrane morphological changes, leading to outer membrane bending and promoting the formation of OMVs[56]. PG constitutes a common component of the cell wall in both G+/G- bacteria[57]. Up-regulating the expression of PG hydrolase PGase enhances PG hydrolysis, thereby inducing the BEVs secretion. This universal strategy has been validated in both G+/G- bacteria. An autonomous PGase expression system yields a 9.37-fold increase in Escherichia coli Nissle 1917 (EcN)-derived BEVs[58]. Furthermore, a combined strategy integrating membrane curvature engineering with dynamic PGase modulation results in a remarkable 149-fold increase in vesicle production[59].
Aside from gene up-regulating modification, another widely utilized genetic engineering method is the knockout/inhibition of genes associated with BEVs formation. This strategy targets the molecules related to crosslinking of the three layers (outer membrane, PG, and inner membrane) of G- bacteria[53,54]. Knockout of OmpA in Acinetobacter baumannii[60] and lipoprotein (Lpp) in E. coli[61], as well as the deletion of nlpI in
Enhancing the production of BEVs through genetic engineering offers a key advantage in achieving fundamental breakthroughs in yield while simultaneously endowing BEVs with new functionalities. However, this strategy also faces significant challenges, including potential growth impairment in modified cells, technical complexity, and stringent regulatory approval requirements when used as pharmaceutical products. Overall, it represents a cutting-edge strategy with immense potential but demanding technical and human safety requirements.
Fermentation optimization
By altering the culture conditions and applying environmental insults, the general stress response of bacteria is induced, thereby increasing the release of BEVs as a protective response[67]. Fermentation optimization mainly involves altering certain nutrients and cultivation parameters.
The lack of Mg2+ increases the protein level of PagL, which catalyzes the deacetylation of lipid A on LPS, and causes membrane bending and release of BEVs, with 3-5-fold increase[67]. The depletion of sulfate and iron from the culture medium can decrease the expression of VacJ/Yrb transporter and lead to phospholipid accumulation, which disrupts the membrane symmetry[65,68]. Also, in Neisseria meningitidis, exhaustion of cysteine and high dissolved oxygen levels cause oxidative stress, which triggers hypervesiculation[69,70].
Besides the change in culture medium components, extreme environmental factors (temperature, pH, ultraviolet light) can also influence the generation of BEVs. In Pseudomonas, when cultivated at elevated temperature, the combined effects of increased membrane fluidity and enhanced interaction of the LPS with Pseudomonas quinolone signal (PQS) synergistically promotes the biogenesis of BEVs[71]. Similarly, during the formation of BEVs in Serratia marcescens, low temperatures of 22 or 30 ℃ induce stress response, leading to the production of large quantities of BEVs[72]. When faced with extreme pH condition and ultraviolet (UV) radiation, the bacteria can develop self-protective mechanisms and generate more BEVs[73,74]. However, the alteration of the culture conditions typically results in BEVs heterogeneity; thus, culture parameter optimization must be combined with rigorous quality assessment.
Physical and chemical stimulation
To enhance BEVs production, two additional strategies can be employed: physical modification and chemical treatment. These methods effectively promote vesicle formation and release by altering the physiological state of bacteria or their membrane stability.
Physical treatment primarily induces BEVs formation through mechanical force. Sonication can enhance BEVs generation by increasing the membrane instability; however, the complete protein repertoire may not be fully retained in the resulting BEVs[75,76]. Nitrogen cavitation generates a mechanical force within a chamber that rapidly disrupts cell membranes, subsequently releasing intracellular contents. The cell membranes spontaneously form nanoscale compartments, producing membrane-derived nanovesicles composed entirely of the original cell membrane, thereby achieving rapid BEVs preparation (up to 20-to 30-fold relative to native secretion) and better preserving protein integrity than sonication[77].
Chemical methods directly target bacterial cell membranes with chemical reagents or induce stress responses to promote BEVs generation. Detergents, such as deoxycholate and sodium dodecyl sulfate, effectively remove LPS from BEVs and destabilize cell membranes to stimulate BEVs secretion while reducing the unintended innate immune response triggered by LPS[56,75,78]. Chelating agents such as Ethylenediaminetetraacetic Acid (EDTA) cause mild membrane instability by neutralizing the negative charge of LPS, an effect that can preserve LPS and natural cargo in BEVs[79-81]. Moreover, exposure to sublethal concentrations of antibiotics (such as ciprofloxacin) can activate protective stress responses in bacteria, promoting BEVs release with 2-to 4-fold enhancement by temporarily altering membrane state[82-84]. Antimicrobial peptides exhibit antibiotic-like effects in inducing BEVs release, but the generated vesicles typically contain higher levels of phosphatidylglycerol, potentially compromising their thermal stability[85,86].
Notably, basal BEV yield varies substantially among bacterial strains. Genetic backgrounds, especially envelope stress response systems and PG network rigidity, can directly influence both native vesiculation levels and the fold‑increase achievable by hypervesiculating mutations, as well as the final product quality. Therefore, strain selection is a prerequisite for achieving target yield and cargo integrity for any given therapeutic application. Moreover, the choice of yield enhancement strategy has notable implications for BEVs quality and scalability. While genetic engineering offers the highest yield improvements and high batch-to-batch consistency, engineered strains may alter size distribution and cargo composition. Regarding scalability, genetically engineered strains are well-suited for industrial fermentation, as the modified bacteria can be cultured at large scale using standard bioreactors. In contrast, physical methods require specialized equipment and are more challenging to scale up, while chemical induction offers simplicity but raises concerns about residual reagent contamination. Therefore, a comprehensive evaluation of the characteristics of the resulting BEVs must be conducted based on specific application objectives.
Engineering the membrane of BEVs
Modifying the membrane surface of BEVs endows them with enhanced targeting and delivery capabilities, improves biosafety and regulates immunogenicity, and optimizes pharmacokinetics and stability. Membrane surface modification methods are primarily categorized into three types: genetic engineering, chemical modifications, and membrane hybridization [Figure 4].
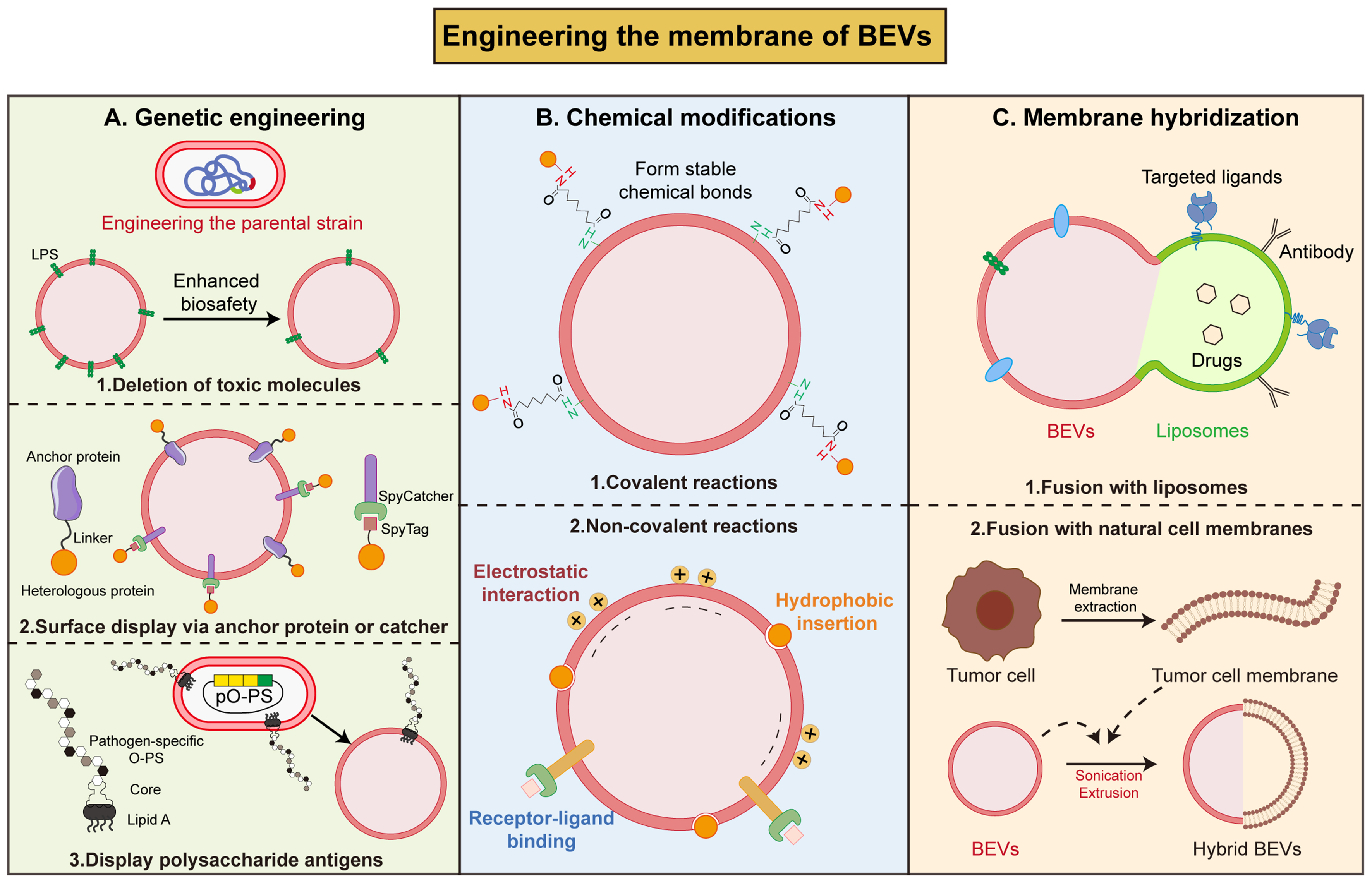
Figure 4. Strategies to engineer the membrane surface of BEVs. (A) Utilizing genetic engineering techniques to eliminate toxic molecules in BEVs membrane surface, fuse desired proteins with anchor protein or catcher, or displaying polysaccharide antigens on BEVs surface; (B) Using covalent (forming stable chemical bonds) or non-covalent (electrostatic interaction, hydrophobic insertion, receptor-ligand binding) reactions to coat antigens on BEVs surface; (C) Fusion with liposomes or natural cell membranes forms hybrid BEVs. BEVs: Bacterial extracellular vesicles; LPS: lipopolysaccharide; SpyTag: SpyTag peptide; SpyCatcher: SpyCatcher protein; O-PS: O-polysaccharide (O-antigen); PO-PS: pathogen-specific O-polysaccharide.
Genetic engineering
Genetic manipulation of parental bacteria at the molecular level enables the deletion of toxic molecules during BEVs formation or the natural expression of desired proteins on the surface during BEVs biosynthesis. Both G+ and G– bacteria can produce molecules with toxic or potentially toxic effects that play important roles in bacterial infections[87]. Knocking out or mutating selected genes in the LPS, TA and LTA synthetic pathways can fundamentally alter the membrane composition of BEVs, in a way that reduces their toxicity[53]. For example, eliminating bacterial virulence genes (msbB) at source via CRISPR/Cas9 gene editing can enhance the biosafety of BEVs[88].
In addition to the engineering of BEVs membrane composition to reduce toxicity, genetic engineering techniques have also been employed to display exogenous proteins on BEVs surface. Through surface display technology, desired functional proteins or peptides are fused with the native membrane proteins of BEVs, enabling the protein to be efficiently and stably anchored to the membrane[89]. Currently, the most common carrier proteins include bacterial cytolysin A (ClyA), OmpA, Omp W and β-lactamases (Bla)[90-92]. ClyA is a pore-forming toxin capable of traversing the bacterial inner membrane to localize to the outer membrane[93]. By fusing exogenous proteins to the C-terminal of ClyA, these proteins can be directly localized to the membrane surface and to the lumen of OMVs[94,95]. Genetic engineering of BEVs by surface display technology is widely used to enhance the immunogenicity[53]. It was reported that the fusion of the coding region of the programmed death-ligand 1 (PD-L1) with ClyA could yield engineered OMVs carrying PD-L1. This genetic modification of OMVs enhances their accumulation at tumor site, and induces blockade of death-ligand 1, resulting in a more effective anti-tumor immune response[96]. Moreover, the fusion of ClyA with cell-penetrating peptide (CPP) achieves an augmented tumor-targeted binding and penetration[97], and fusion with hyaluronidase (HAase) degrades the extracellular matrix (ECM) within the tumor microenvironment, thereby enhancing the delivery efficiency of therapeutic drugs[98]. Beyond the ClyA system, autotransporter-based platforms have also been developed for antigen display on membrane surfaces. The hemoglobin protease (Hbp) autotransporter from E. coli has been engineered into a versatile surface display carrier that can be expressed at high density on OMVs. By fusing heterologous antigens to Hbp, multiple Mycobacterium tuberculosis antigens (ESAT6, Ag85B, and Rv2660c) were successfully displayed on the surface of OMVs, eliciting antigen-specific immune response[99]. As for G+ bacteria (Bacillus subtilis, B. subtilis), Chen et al. have constructed a peptide display platform by fusing spore coat protein CotC with antimicrobial peptide Cathelicidin-BF (Cathe). The BEVs secreted by this engineered strain contains Cathe and demonstrates antimicrobial activity and further alleviates dextran sulfate sodium (DSS)-induced colitis[100].
Apart from protein/peptide components, utilizing glycoengineering strategies, polysaccharide antigens can be inserted on the BEVs surface to generate glycoengineered BEVs as vaccine candidates[101]. Price et al. developed a modular plasmid containing crucial genes for enzymes of S. pneumoniae serotype 14 capsule (CPS14) synthesis. When this plasmid was transformed into O-antigen-deficient E. coli, the synthesized capsular polysaccharide could bind to the lipid A core of LPS, thereby enabling the generation of eBEVs carrying the CPS14 antigen to prevent multiple infections[102]. Similarly, expression of tularemia O antigen polysaccharide (O-PS) in O-antigen-deficient E. coli enables the assembly of glycosylated BEVs with protective immunogenic activity via the core glycosylation of LPS[103].
The core advantages of utilizing genetic engineering for BEVs membrane modification lie in its precision and in situ nature. By modifying the parental bacterial genome, the modifying components can be displayed
Chemical modifications
Employing biochemical reactions, exogenous functional molecules are attached to the surface of BEVs, endowing them with enhanced targeting capabilities and therapeutic potential. Chemical methods are mainly categorized into covalent and non-covalent binding.
Covalent reaction anchors exogenous molecules to the membrane surface via the formation of stable chemical bonds, including conjugation, click chemistry, and aldehyde amine condensation to modify BEVs[104]. Forming amide bonds between the carboxyl groups present on the surface of BEVs and the amino groups of 3-aminophenylboronic acid (PBA) allows the attachment of PBA to the surface of BEVs. The chemically modified BEVs possesses the ability to target sialic acids present on the surface of tumor cells. Furthermore, HAase expression combined with PBA surface modification increased deep tumor penetration of OMVs by 266.7% and enhanced small interfering RNA (siRNA) silencing efficiency by 70.5%[105]. In addition, the use of cross-linking agents such as thiol-maleimide, adipic acid, or carbodiimide makes it possible to covalently attach protein antigens, inhibitors or nanobodies to BEVs[106,107]. To streamline the process, researchers have developed improved methods based on bis(sulfosuccinimidyl)suberate cross-linking or reductive amination, enabling direct coupling via lysine residues on the antigen without requiring pre-derivatization[107,108]. This feature renders the Generalized Modules for Membrane Antigen (GMMA) system an efficient “plug-and-play” vaccine development platform.
Non-covalent modifications rely on diverse physical interactions and specific-binding, offering a flexible and efficient alternative approach for the functionalization of BEVs[104]. Firstly, molecular attachment can be realized through multivalent electrostatic interactions or hydrophobic insertion. Sun et al. modified Angiopep-2 and Trans-Activator of Transcription (TAT) peptide onto the surface of BEVs membrane via lipid insertion, enabling the encapsulation of a chemotherapy drug[88]. The SpyCatcher-SpyTag protein conjugation system enables efficient, modular antigen display on BEVs surfaces by forming specific isopeptide bonds with SpyTag peptides on target proteins[109]. Furthermore, through receptor-ligand binding, RGD peptide binds to integrin αvβ3 for the loading of indocyanine green[110]. These non-covalent approaches collectively broaden the applications of BEVs in drug delivery and immunotherapy.
Chemical modification offers diverse strategies for functionalizing BEVs: covalent modifications provide stable, durable linkages suitable for vaccine development and targeted therapies; non-covalent modifications offer greater flexibility for drug delivery, molecular display, and rapid functionalization. Collectively, these approaches advance the application prospects of BEVs in drug delivery, immunotherapy, and diagnostics.
Membrane hybridization
Through physical forces (extrusion, ultrasonication, freeze-thaw cycles), BEVs can be fused with functional liposomes or cell membranes to form hybrid vesicles that combine the characteristics of both. The emerging EVs-liposome fusion strategy has developed into a powerful approach to overcome the limitations of EVs in application, and expand the functional capabilities of EVs[111]. As liposomes are artificially synthesized, their surfaces can be modified to achieve targeted, prolonged, and sustained release, while possessing advantages as high biocompatibility and low immunogenicity[112]. Zhai et al. engineered hybrid vesicles by fusing OMVs derived from attenuated Salmonella with photothermal sensitive liposomes, demonstrating significant antitumor efficacy[113]. Also, Pan et al. used modified liposomes loaded with luteolin and borneol to fuse with OMVs isolated from Stenotrophomonas maltophilia using an extrusion technique. The constructed hybrid OMVs-liposomes achieve deep mucosal penetration capability and retention capacity within the inflamed colon[114]. Integrating liposomes technology with BEVs produces innovative nano-pharmaceuticals that simultaneously exploit the immunogenicity of BEVs and the precise targeting and tunable drug release profiles of liposomes, paving the way for a versatile platform in targeted therapy.
Another strategy for BEVs membrane hybridization is the fusion with natural cell membranes, especially eukaryotic membranes. Abundant studies have reported hybrid membrane-based nanovesicles by combining bacterial OMVs with tumor cell membranes to confer the unique advantages of BEVs as personalized cancer vaccines in situations where neoantigens are not readily accessible[115]. Specifically, Wang et al. reported a hybrid membrane platform by fusing OMVs with B16-F10 tumor cell membrane to utilize both the advantages of OMVs and tumor cells to achieve better antitumor efficacy[116]. In another work, researchers showed the successful construction of a heterologous fusion membrane tumor vaccine by fusing BEVs from Staphylococcus aureus with B16-F10 cell membrane to activate immune responses and prevent tumor development[117]. Wang et al. constructed hybrid nanovesicles by fusing OMVs secreted by E. coli with membranes extracted from tumor-homing macrophages to selectively target tumor tissues and inhibit tumor growth[118]. Additionally, extensive research has also been conducted to obtain hybrid BEVs from other bacteria. Firstly, A. muciniphila, Bifidobacterium longum (B. longum), and Bifidobacterium breve (B. breve) were treated with lysozyme to prepare protoplasts. Then, hybrid vesicles were synthesized via ultrasonication and extrusion, which demonstrated enhanced targeting capacity towards tumors and peripheral lymphoid organs[119].
In summary, engineering modifications to the surface of BEVs represent a key strategy for optimizing the
Engineering the lumen of BEVs
By using exogenous (physical and chemical methods) and endogenous (genetic engineering) strategies, therapeutic cargos are loaded into the lumen of BEVs, thereby further enhancing the functions of BEVs [Figure 5].
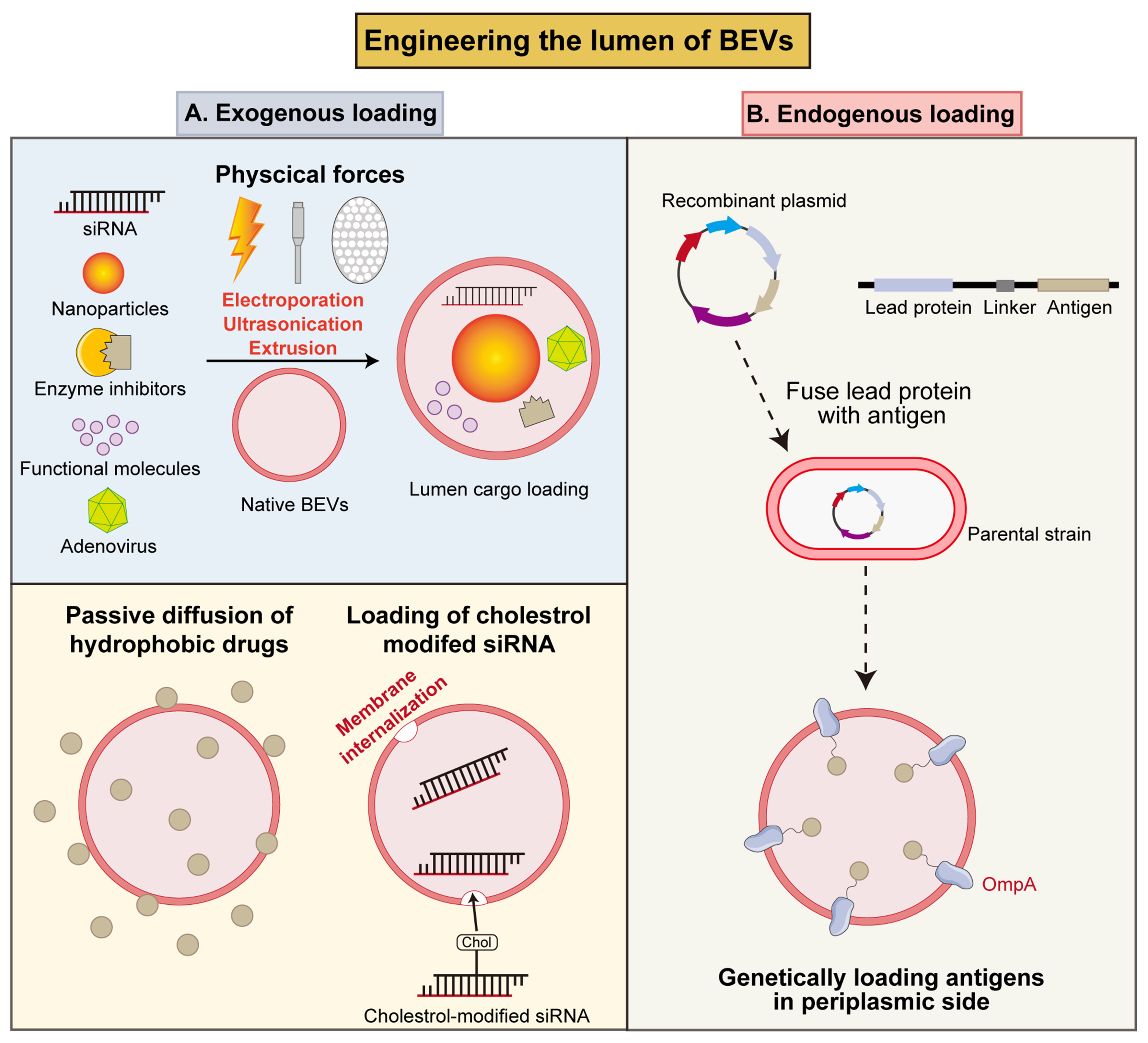
Figure 5. Strategies to engineer the lumen of BEVs. (A) Exogenous loading of cargos by physical forces (electroporation, ultrasonication and extrusion), passive diffusion, or cholesterol modification into BEVs lumen; (B) Endogenous loading of antigens into the BEVs lumen through genetic engineering strategy. BEVs: Bacterial extracellular vesicles; siRNA: small interfering RNA; OmpA: outer membrane protein A.
Exogenous loading
Firstly, physical treatments could achieve the loading of exogenous cargos in the inner cavity of BEVs. Electroporation has been used to load small gold nanoparticles (AuNPs) in Pseudomonas aeruginosa-derived BEVs via the pores in the lipid bilayer generated by a transient electric field[120]. Also, electroporation successfully loads siRNA and specific inhibitors in BEVs to regulate tumor microenvironment and further inhibit the tumor growth[121,122]. Another study utilized ultrasonic bath to load fucoxanthin (Fx) in Lactobacillus plantarum (L. plantarum) derived synthetic membrane vesicles to treat colitis[123]. Ban et al. encapsulated oncolytic adenovirus (Ads) in E. coli derived BEVs through extrusion to enhance the intra-tumoral accumulation of Ads and achieve autophagy-cascade-augmented immunotherapy[124].
Furthermore, chemical approaches represent another prospective nanotechnology for the loading of therapeutic molecules. A prevalent strategy for antibiotic loading into the BEVs lumen is passive loading, which is based on co-culturing bacteria in media containing sublethal concentration of antibiotics, which facilitates the intrinsic incorporation of antibiotics during the natural biogenesis of BEVs[125]. Additionally, cholesterol-modified siRNA can be loaded into BEVs via hydrophobic interactions[105,126]. This mild, non-invasive encapsulation strategy not only achieves efficient siRNA loading, but also preserves the integrity and biological activity of both BEVs and the cargos.
Endogenous loading
As for the development of BEVs vaccine, it is crucial to ensure the high immunogenicity of BEVs[127]; this requires antigen-presenting cells (APCs) to engulf antigen-containing particles. Thus, engineering the parental bacteria to load more antigens offers a method for enhancing immunogenicity by localizing bioactive molecules within the periplasmic space[53]. Multiple antigens were successfully encapsulated into the lumen of BEVs, thereby inducing protective immune responses against pathogen infection. For instance, Chlamydia muridarum/Chlamydia trachomatis serovar D (DO) DO serine protease HtrA antigen can induce severe humoral and cellular immune responses in vivo. In one report, HtrA was fused with OmpA and expressed in E. coli to deliver it to the lumen of BEVs. The obtained eBEVs effectively stimulate functional immune responses[128]. Similarly, Fantappiè et al. selected a group of heterologous antigens (GAS Slo, SpyCEP, Spy0269 and GBS SAM_1372) to fuse with OmpA. The antigens incorporated into the BEVs lumen induced the production of high antibody titers in mice[129]. Collectively, these results indicate that BEVs carrying internalized antigens undergo structural alterations upon interaction with immune cells, leading to antigen exposure or release, which is then recognized by ACPs to mount a specific antibody response. Thus, the simplicity of genetic engineering paves the way for diverse strategies to load antigens into BEVs.
Comparative summary of engineering strategies
The engineering strategies discussed above can be broadly categorized into three main technologies: genetic engineering, physical methods, and chemical approaches, each with distinct advantages, limitations, and application scenarios.
Genetic engineering enables precise yield enhancement, surface display, and LPS detoxification with high scalability but faces technical complexity and regulatory hurdles. Physical methods (sonication, cavitation, extrusion) are rapid and broadly applicable but risk cargo damage and batch inconsistency, issues that need to be solved. Chemical approaches offer operational simplicity and versatile functionalization but may compromise BEV integrity or leave residues. The optimal strategy depends on the application, bacterial strain, and regulatory pathway.
The choice of engineering strategy ultimately depends on the intended application, the bacterial strain involved, and the regulatory pathway. Often, hybrid strategies combining two or more approaches yield the most clinically promising BEV platforms. Table 2 provides a systematic comparison of these strategies, highlighting their relative strengths and limitations, as well as the typical applications.
Comprehensive comparison of different BEVs engineering strategies
| Strategies | Specific approaches | Advantages | Disadvantages | Applications | Ref. |
| Genetic engineering | Gene knockout/overexpression (nlpl, ompA, lpp, ompT, pagL, degS, degP) | 1. Significant increase in BEVs yield; 2. Uniform and stable BEVs composition; 3. Suitable for large-scale production; 4. Endow BEVs with inherent functions | 1. High technical requirements; 2. Time-consuming; 3. May affect the growth and viability of the engineered strain | Mass production of standardized BEVs; Vaccine development | [55,56,60-64] |
| Surface display system (ClyA, OmpA, Lpp-OmpA) | 1. In situ, covalent, high-density display; 2. Excellent stability | 1. Fusion proteins may affect the function of anchor proteins; 2. Strain-dependent efficiency | Targeted ligand presentation; Vaccine antigen delivery | [89-98] | |
| LPS/Lipid A modification (msbB, lpxL, lpxM, pagL, pagP) | 1. Reduce endotoxin toxicity at source; 2. Preserve natural adjuvant activity | 1. Excessive attenuation may reduce the immunostimulatory effect; 2. The strategies for modifying different bacteria vary | Therapeutic applications with high safety requirements; Development of vaccine adjuvants | [88] | |
| Physical methods | Ultrasonication, nitrogen cavitation, extrusion | 1. Rapid and straightforward; 2. Do not require genetic manipulation; 3. Applicable to multiple strains | 1. May compromise the natural structure of the cargo; 2. Poor batch-to-batch consistency; 3. Risk of contamination by impurities; 4. High dependence on equipment | Rapid preparation of BEVs; Strains that are difficult to genetically modify; Preparation of hybrid vesicles | [75-77,124] |
| Membrane fusion | 1. Combine the immunogenicity of BEVs with the targeted/controlled-release properties of synthetic carriers; 2. High degree of functional designability | 1. Unstable fusion efficiency; 2. High heterogeneity of fusion products; 3. Complex quality control | Targeted drug delivery; Personalized cancer vaccines | [113-118] | |
| Chemical approaches | Surface covalent modification (amide bonds, click chemistry, cross-linking agents) | 1. Stable and long-lasting connections; 2. Controllable level of functionality; 3. High “plug-and-play” flexibility | 1. Affect the function of natural proteins on the surface of BEVs; 2. Residual reagents pose a safety risk | Vaccine development (GMMA platform); Targeted ligand conjugation | [104-108] |
| Surface non-covalent modification (electrostatic adsorption, hydrophobic insertion, ligand-receptor binding) | 1. Gentle operation and simple conditions; 2. Does not damage the structure of BEVs; 3. High functionalization efficiency | 1. The stability of the modification is relatively poor | Rapid functionalization screening; Development of drug delivery carriers | [88,105,109,110] | |
| Chemically induced hypersecretion (detergents, EDTA, antibiotics) | 1. Easy to operate and cost-effective; 2. Significant increase in yield; 3. Simultaneously reduces LPS content | 1. Residual reagents may compromise safety; 2. Alter the composition and immunogenicity of BEVs; 3. Poor batch-to-batch consistency | Rapid expansion of laboratory capacity; Preparation of attenuated BEVs | [56,75,78-84] |
THERAPEUTIC APPLICATION OF eBEVs
Leveraging the versatile engineering strategies described above, eBEVs have been developed into tailored therapeutic platforms for a wide range of diseases. Compared with nBEVs, eBEVs offer enhanced targeting accuracy, improved cargo delivery efficiency, and tunable immunomodulatory properties. To date, preclinical studies have demonstrated the promising potential of eBEVs in treating intestinal disorders, metabolic diseases, autoimmune conditions, cancers, central nervous system (CNS) disorders, and skin injuries. In this section, we systematically review the application of eBEVs across these disease categories [Figure 6].
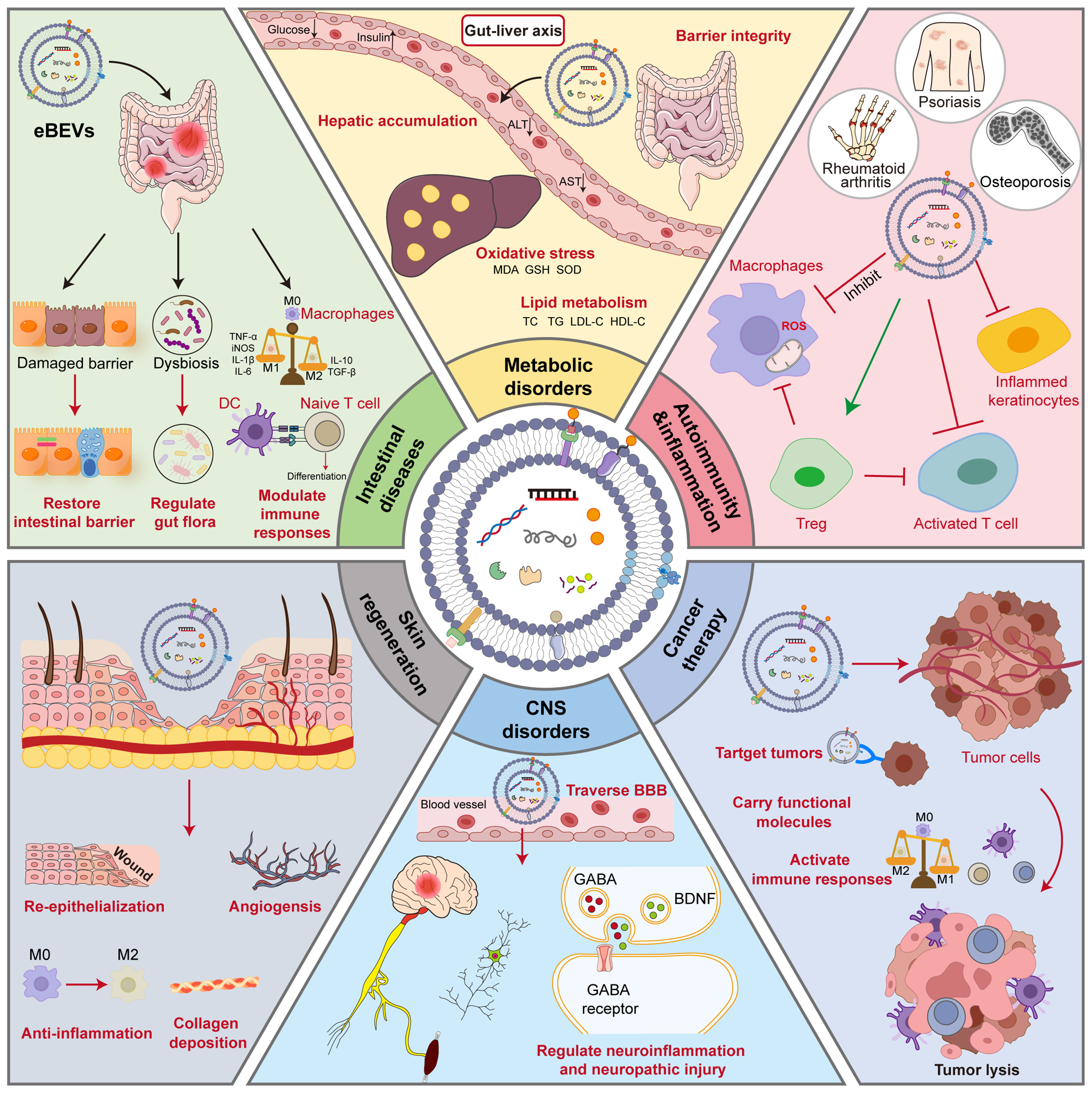
Figure 6. Application of eBEVs in treating various diseases. eBEVs could participate in disease regulation through multiple pathways and targets, covering aspects such as intestinal barrier repair, immune regulation, metabolic intervention, neuroprotection, and anti-tumor activity. Firstly, eBEVs can act on damaged intestinal barriers by restoring their integrity and regulating the balance of the microbiota. In metabolic disorders, eBEVs participate in regulating oxidative stress indicators (such as MDA, GSH, SOD) and lipid metabolism-related indicators (TC, TG, LDL-C, HDL-C), thereby intervening in metabolic disorders. In autoimmune diseases such as psoriasis, rheumatoid arthritis, and osteoporosis, eBEVs demonstrate regulatory potential. eBEVs can regulate the differentiation of naive T cells, affect immune responses and inflammasome activity, promote the function of Treg, and regulate the transformation of macrophages from M1 type to M2 type, exerting anti-inflammatory effects. In tumor treatment, eBEVs can carry functional molecules, activate immune responses, and directly promote tumor cell lysis, demonstrating targeted therapeutic potential. For central nervous system diseases, eBEVs can cross the blood-brain barrier and regulate GABA, BDNF, and related receptor pathways, thereby alleviating neuroinflammation and nerve damage. eBEVs can also promote skin regeneration by promoting re-epithelialization, vascular regeneration, and collagen deposition. This figure reflects the diversity and synergy of eBEVs as multifunctional nanocarrier, highlighting the integrated mechanism in barrier repair, immune remodeling, metabolic regulation, neuroprotection, and tumor intervention. eBEVs: Engineered extracellular vesicles; BEVs: bioengineered extracellular vesicles; ALT: alanine aminotransferase; AST: aspartate aminotransferase; MDA: malondialdehyde; GSH: glutathione; SOD: superoxide dismutase; TC: total cholesterol; TG: triglycerides; LDL-C: low-density lipoprotein cholesterol; HDL-C: high-density lipoprotein cholesterol; ROS: reactive oxygen species; TNF-α: tumor necrosis factor-α; iNOS: inducible nitric oxide synthase; IL-1β: interleukin-1β; IL-6: interleukin-6; TGF-β: transforming growth factor-β; DC: dendritic cell; Treg: regulatory T cell; BBB: blood-brain barrier; CNS: central nervous system; GABA: γ-aminobutyric acid; BDNF: brain-derived neurotrophic factor.
Intestinal diseases
nBEVs, acting as pivotal mediators of microbe-host interactions, can regulate intestinal immune homeostasis and barrier function through synergistic effects. The mechanism can be summarized in the following aspects; first, nBEVs can directly enhance the integrity of the intestinal epithelial barrier. For instance, Roseburia intestinalis-derived BEVs are reported to upregulate the expression of tight junction proteins, thereby reducing intestinal epithelial permeability and thus collaborating with the host to resist pathogen invasion[130]. Similarly, L. plantarum and its BEVs can also promote mucus secretion to preserve and repair the gut barrier[131]. Secondly, nBEVs can modulate the phenotypic polarization of innate immune cells, particularly macrophages. BEVs derived from L. plantarum and L. johnsonii can induce macrophages to polarize towards the anti-inflammatory M2 phenotype[131,132]. In addition, nBEVs can reshape the gut microbiota composition by increasing the abundance of beneficial bacteria and suppressing the abundance of potential pathogenic bacteria, thus restoring the microbial balance[47]. Collectively, nBEVs constitute a multidimensional therapeutic network through a synergistic mechanism of “barrier repair-immunomodulation-microbiome remodeling”, offering a potential intervention strategy for gastrointestinal diseases like colitis.
eBEVs, as an emerging nanotherapeutic platform, demonstrate enormous potential in the treatment of intestinal diseases owing to their inherent biocompatibility, superior mucosal penetration capabilities, and amenability to engineering modification. Through targeted modification and functional enhancement of nBEVs, the resulting eBEVs demonstrates high targeting precision and synergistic therapeutic effects. Because the production of nBEVs is typically low, researchers have genetically engineered E. coli and
Engineered modifications systematically address the limitations of nBEVs in drug loading capacity, targeting precision, and controllable efficacy through multidimensional strategies. These intelligent eBEVs systems not only amplify their inherent capabilities to repair the gut barrier and modulate the microbiota composition, but also integrate precise drug delivery with active immune modulation in intestinal diseases.
Metabolic disorders
Metabolic disorders, such as type 2 diabetes mellitus (T2DM), obesity, metabolic-associated fatty liver disease (MAFLD), and hyperuricemia, have become critical global public health challenges[139-141]. Conventional drug therapies exhibit limitations in targeting specificity, bioavailability, and multi-target regulation. Recently, EVs have emerged as novel nanocarriers, demonstrating significant potential in the intervention of metabolic disorders due to their inherent biocompatibility, low immunogenicity, and exceptional ability to penetrate biological barriers[142].
Mammalian EVs have demonstrated therapeutic potential. For example, human umbilical cord mesenchymal stem cell-derived EVs enriched with Exendin-4 directly improve glycemic control and renal function in diabetic nephropathy[143]. Besides, plant-derived exosome-like nanovesicles (PELNs) from various sources also ameliorate metabolic disorders through multiple pathways. Pomegranate-derived exosome-like nanovesicles (naturally rich in ellagic acid) are reported to restore intestinal barrier function and reduce liver oxidative stress and fibrosis markers in MAFLD model[144]. Ginger-derived exosome-like nanoparticles can activate the Phosphatidylinositol 3-kinase/Protein Kinase B (PI3K/Akt) signaling pathway in the liver via their carried miRNAs, which further enhances insulin sensitivity, inhibits gluconeogenesis, and produces hypoglycemic effects in T2DM mice[145]. Similarly, the bitter melon-derived EVs loaded with curcumin demonstrate unique targeted delivery capabilities, enabling specific transport of active components to the pancreas via the lymphatic system to restore insulin secretion function[146]. Despite the remarkable therapeutic efficacy of these naturally sourced EVs, their complex active constituents and inconsistent batch-to-batch variability have hindered their applications, which has propelled the field towards the development of eBEVs.
The core concept of eBEVs involves transforming probiotics and other GRAS (Generally Recognized As Safe) microorganisms into “living factories” that continuously produce and load therapeutically active proteins into BEVs. This approach addresses the global challenge of delivering macromolecular biological agents (such as enzymes and peptides) via oral administration. The utilization of microfluidics-assisted ultrasound technology allows the encapsulation of Fx within Lactobacillus-derived EVs (LCEV), forming LCEV@Fx. BEVs encapsulation prolongs Fx retention time within the GI tract, enhances absorption across the entire serosal layer, and increases its hepatic accumulation. LCEV@Fx reduced white fat accumulation and ameliorated hepatic steatosis in high-fat diet mice by promoting fatty acid oxidation via the AMP-activated protein kinase/Sirtuins 1 (AMPK/SIRT1) pathway and downregulating lipid synthesis-related proteins[147]. In another landmark study, researchers genetically engineered probiotics to produce high yields of OMVs, enabling the targeted therapeutic protein (uric acid oxidase) to self-assemble within the OMVs with an encapsulation rate as high as 97.9%. OMV encapsulation provides effective protection for unstable protein drugs, allowing them to withstand gastric acid and digestive enzymes while efficiently traversing the intestinal epithelial barrier into systemic circulation. In hyperuricemic model, oral administration of such eBEVs resulted in continuous clearance of uric acid[63]. These techniques are adaptable to multiple therapeutic proteases, providing a universal platform for intervention in a range of metabolic disorders.
The intervention of metabolic diseases with BEVs is in its early stages. Although BEVs protect protein cargo from gastric acid and proteases, efficient intestinal absorption remains challenging. BEVs must traverse the mucus layer, avoid entrapment by intestinal epithelial glycocalyx, and undergo transcytosis to reach systemic circulation. Protein release may occur prematurely in endosomes, reducing bioavailability. Furthermore, protein loading efficiency and maintaining native protein folding during encapsulation are additional hurdles. Future engineering should focus on muco-adhesive surface modifications and endosomal escape enhancement.
Autoimmunity and inflammation
Notably, EVs from different sources have also demonstrated tremendous potential in autoimmune and inflammation-related diseases, such as rheumatoid arthritis, psoriasis, and osteoporosis, exerting therapeutic effects through multiple mechanisms including remodeling the immune microenvironment, regulating inflammatory responses, and promoting tissue repair.
Mammalian EVs and PELNs are the candidates to be explored in the treatment of autoimmune and inflammation-related diseases. Mammalian EVs possess strong ability in regulating innate immune responses and tissue repair signals[148]. Recently, intestinal organoid-derived EVs are reported to regulate the gut-bone axis, further exerting therapeutic functions for osteoporosis[149]. PELNs, as edible nanocarriers, demonstrate increasingly evident therapeutic potential in this area[150,151]. A typical example is ginger-derived EVs (GDEVs), which are naturally rich in anti-inflammatory components such as 6-gingerol. Researchers engineered folate-modified GDEVs (FA-GDEVs) to enhance the targeting ability of GDEVs. Folate receptors are highly expressed on pro-inflammatory M1 macrophages within rheumatoid arthritis joints, enabling FA-GDEVs to actively target inflammatory sites. FA-GDEVs can effectively downregulate pro-inflammatory factors such as TNF-α and IL-6, thus significantly reducing joint swelling and bone erosion[150,151]. Despite advances in mammalian and plant-derived EVs, challenges still persist, prompting researchers to turn attention towards BEVs, which offer greater programmability and potential for mass production.
Probiotic-derived EVs, as postbiotics, exert their effects in rheumatoid arthritis through multiple pathways, including modulating immune cells, reducing pro-inflammatory factors, restoring intestinal barrier integrity, and inhibiting osteoclast activity[152]. For diseases requiring multi-stage sequential treatment, such as osteoporotic fractures (OPF), researchers have developed sophisticated BEV-based platforms that address the distinct phases of healing. Building on prior work demonstrating the feasibility of synthetic biology-based BEVs for bone-related conditions[153], a recent breakthrough study used synthetic biology techniques to engineer bacteria that produce eBEVs simultaneously carrying bone morphogenetic protein-2 (BMP-2) and vascular endothelial growth factor (VEGF). Subsequently, the constructed BEVs carrying BMP‑2 and VEGF (BEVs‑BP)were co-encapsulated with IL-4 within a methacrylamide-modified gelatin (GelMA) hydrogel, forming IL-4/BEVs-BP@GelMA. This system enabled temporally controlled release: upon injection into the fracture site, the hydrogel rapidly released IL-4, swiftly ameliorating the inflammatory microenvironment at the lesion site. Then, the BEVs-BP were slowly released and, after internalization by surrounding cells, the secretion of BMP-2 and VEGF promoted osteogenic differentiation and angiogenesis[154]. As for the treatment of psoriasis, a novel strategy has been developed that consists of the large-scale production of
Therapies based on eBEVs hold promise for delivering safer, more effective and personalized treatment options for patients with autoimmune and inflammatory diseases, thereby genuinely driving transformative change in clinical treatment paradigms.
Cancer therapy
In the field of cancer therapy, BEVs, particularly OMVs, have emerged as a highly promising next-generation smart nanomedicine platform. This is due to their innate immune-activating properties, exceptional tumor tissue enrichment capacity [the enhanced permeability and retention effect (EPR effect)], and high degree of engineering potential[156]. Other core advantages of the use of OMVs reside in the integration of multiple functions: targeted delivery, immune activation, and drug loading. Currently, eBEVs for cancer therapy primarily focus on four key strategies: surface functionalization and modification, therapeutic payload delivery, in situ construction of living factories, and regulation of the immune microenvironment.
Surface functionalization modifications aim to enhance the targeting efficiency and improve the safety profile of nBEVs, mainly through genetically engineered expression of fusion proteins, modification of target peptides, and fusion of hybrid membranes. eBEVs are uniquely positioned to address tumor heterogeneity. Their surface display platforms allow simultaneous presentation of multiple tumor-associated antigens, broadening the repertoire of targeted clones. For instance, researchers reported the use of genetic engineering techniques to express the vesicular stomatitis virus G protein on the surface of OMVs, enabling them to fuse with tumor cell membranes and achieve tumor heterogenization. These bacterial components can enhance the adjuvanticity and antigenicity of heterogenized tumor cells, effectively mobilizing the innate immune system to recognize and phagocytose tumors[157]. Furthermore, through techniques such as co-culture, electroporation, and chemical loading, therapeutic agents including chemotherapy drugs and nucleic acids can be encapsulated within BEVs, enabling the precise delivery of therapeutic payloads to tumor sites. Moreover, BEVs can serve as synthetic biologically-driven in vivo therapeutic factories, thus addressing the complexities of ex vivo production, purification, and drug loading processes. Sun et al. reported the administration of a genetically engineered E. coli, which produces high-yield OMVs loaded with therapeutic RNA, to mice via oral or injectable routes. The bacteria subsequently colonized tumors and continuously produced self-assembled OMVs loaded with PD-L1 siRNA within the tumor microenvironment, with relative RNA abundance 104-107 times higher than the mock group, leading to around 2-fold downregulation of PD-L1 gene expression and about 49.37% tumor suppression, achieving
Lastly, numerous studies have directly eBEVs into cancer vaccines or immunomodulators, achieving regulation of the tumor immune microenvironment through mechanisms such as presenting tumor antigens and loading immunological adjuvants[159]. These distinct modification strategies are often employed in combination to enhance the application of BEVs in cancer therapy. A typical example consists of the genetic modification of OMVs derived from engineered low endotoxicity E. coli and functionalized with peptide angiopep-2 to obtain a nanoplatform capable of continuously traversing the blood-brain barrier (BBB) and targeting glioblastoma. Additionally, this platform is further loaded with doxorubicin and CD47 siRNA. Upon tumor targeting, the immunogenic components of the OMVs synergize with the encapsulated drugs to reprogram tumor-associated macrophages and microglia, effectively overcoming the immunosuppressive microenvironment of brain tumors[160]. eBEVs have also demonstrated remarkable synergy with immune checkpoint blockade therapy. One representative strategy involves genetically engineering OMVs to display the PD-1 ectodomain (OMV-PD1), which simultaneously activates systemic anti-tumor immunity via OMV’s intrinsic immunogenicity and blocks the PD-1/PD-L1 axis, resulting in superior tumor growth inhibition compared to either therapy alone[96].
Compared to lipid or synthetic polymer nanoparticles, eBEVs possess intrinsic self-adjuvant properties due to their abundant pathogen-associated molecular patterns (PAMPs), which activate innate immunity without requiring external adjuvants, while offering a superior safety profile. Strain selection also determines the intrinsic immunogenicity of eBEVs. The LPS‑rich OMVs from S. typhimurium provide potent adjuvant effects, whereas the milder, LPS‑free CMVs from Lactobacillus deliver safer, low‑immunogenicity vehicles for targeted drug delivery. Taken together, eBEVs have evolved from simple bacterial nanostructures into functionally programmable antitumor agents through the convergence of synthetic biology and nanotechnology. Future developments will focus on multifunctional integration, precise controlled release, and addressing standardization and safety challenges in large-scale production and clinical translation. As research progresses, eBEVs hold promise to pioneer novel technical pathways for precision immunotherapy and combination cancer therapies.
CNS disorders
CNS disorders, which affect the brain and spinal cord, constitute a complex and life-threatening category of diseases. Due to their progressive nature and limited treatment options, they present a significant global health challenge[161]. While the BBB maintains CNS stability, it also blocks over 98% of potential therapeutic compounds, presenting a major obstacle to effective treatment[162]. Developing effective therapies for CNS disorders requires overcoming two primary challenges: crossing the BBB and addressing complex underlying pathological mechanisms.
EVs from diverse sources, acting as nanoscale messengers, have been demonstrated to traverse the BBB through various pathways, making them a promising therapeutic avenue for treating CNS diseases[163]. Mesenchymal stem cell-derived exosomes (MSC-Exos) can enhance functional recovery in stroke rats while promoting angiogenesis, neurite remodeling and neurogenesis[164]. Also, MSC-Exos can reduce inflammation and inhibit abnormal neurogenesis on astrocytes[165]. PELNs, acting as neuroprotective agents, can modulate pathological processes such as neuroinflammation and oxidative stress by delivering special plant-derived bioactive molecules, including microRNAs, lipids and polyphenols[166,167]. Currently, an increasing number of studies are turning their attention to BEVs, owing to their close association with the gut-brain axis and their amenability to engineering modifications[22].
BEVs can transmit crucial signals via the gut-brain axis[168], thereby regulating neuroinflammation and neuropathic injury. L. paracasei-derived EVs show the ability to alleviate hyper-ammonemic rat models by reducing neuroinflammation, restoring GABA neurotransmission, and improving motor coordination by mediating “gut-brain axis” communication[169]. Moreover, BEVs may also improve cognitive deficits by regulating neurotrophic factors such as brain-derived neurotrophic factor (BDNF) and other proteins related to cognitive function[170]. However, research reports on the direct application of eBEVs in the treatment of CNS diseases remain exceedingly scarce. Nevertheless, portable engineering strategies from successful cases in other fields can be extracted and applied. Firstly, since the brain has a unique immune-privileged environment, and the surface of BEVs contains pathogen-associated molecular patterns such as LPS[14], they may trigger strong neuroinflammatory responses, which inspires future studies to modify BEVs to achieve “reduced immunogenicity”. Secondly, neurotrophic factors (such as BDNF, GDNF) or siRNAs targeting Aβ/tau can be loaded into BEVs through synthetic biology or surface display targeting peptides.
In summary, the application of eBEVs in CNS disorders remains at the conceptual stage. Compared to other nanocarriers, eBEVs face the additional challenge of PAMP-induced neuroinflammation, as LPS or TA/LTA can trigger microglial activation and exacerbate CNS pathology. Moreover, the heterogeneous size of BEVs may affect their transport across the BBB. Achieving consistent BBB penetration while avoiding off-target brain inflammation remains a key hurdle for eBEV-based CNS therapies. Future breakthroughs may hinge on the combination of different approaches, for example, developing conditional penetration engineering strategies that intelligently respond to the microenvironment of brain lesions. As well, integrating materials science with synthetic biology to construct biomimetic hybrid eBEVs that minimize immunogenicity represent a promising direction.
Skin regeneration
Beyond the aforementioned applications, EVs demonstrate significant potential in promoting tissue regeneration and repair due to their inherent biocompatibility, low immunogenicity, and exceptional capacity as carriers for bioactive molecules. EVs derived from mammalian cells have been extensively studied, with those originating from stem cells or functionally specialized cells demonstrating potent and comprehensive restorative capabilities[171,172]. PELNs naturally harbor active components capable of modulating the functions of diverse cell types, including stem cells and fibroblasts, thereby regulating cellular behavior and influencing key pathophysiological processes involved in wound healing[173].
BEVs offer a novel and more cost-effective pathway in treating skin problems, with non-animal-derived ingredients aligning more closely with consumer expectations. nBEVs, especially those from probiotics, act by delivering intrinsically bioactive molecules such as miRNA. These molecules can promote epithelialization and angiogenesis[174,175], while regulating macrophage mitochondrial function[176], and ultimately facilitating wound repair. The engineering modification approaches primarily work at three core dimensions: functional component loading, exploration of new functional components, and delivery system optimization, which aim to overcome the therapeutic limitations of nBEVs.
Loading functional components represents the most direct enhancement strategy. For example, the incorporation of catalase and growth factors in BEVs can improve the oxidative stress microenvironment at the wound site and also accelerate tissue regeneration. A genetic engineering strategy that localizes superoxide dismutase (SOD) on B. subtilis-derived EVs could greatly enhance the stability and maintain the activity of SOD, and exhibits potential in restoring skin damage[177]. In addition to conventional loading, discovering and exploiting certain special functional molecules carried by BEVs also represents a promising strategy. For instance, it was found that BEVs derived from Streptococcus mutans (Sm EVs) can effectively promote skin healing, by carrying the transfer RNA (tRNA) fragments. By utilizing electroporation to load high amounts of tRNA isolated from Sm EVs into EcN EVs, it was found that the tRNA could activate the Toll-like Receptor 3 (TLR3) pathway, thereby promoting the proliferation and migration of epithelial cells and the healing of skin wounds[178]. Lastly, constructing multi-functional delivery systems, the fusion of Lactobacillus EVs into a photothermal hydrogel created an integrated platform “optical heating-oxygen release-BEVs therapy”. Near-infrared light irradiation can generate a mild thermal effect for antibacterial purposes and promote the release of BEVs. The oxygen released by BEVs synergistically regulates the phenotype of macrophages, achieving a complete set of responses that include antibacterial and anti-inflammatory effects to promote angiogenesis[179]. In another study, microjet technology was used to create artificial LCEV for skin rejuvenation and prevention of photoaging[180]. These examples all demonstrate the potential of eBEVs in the field of cosmetic skincare.
In conclusion, the engineering transformation has enabled BEVs to become programmable “intelligent nanorobots”. Through various engineering transformation strategies, eBEVs demonstrate significant potential to outperform their natural versions in addressing the complex pathological conditions of chronic wounds (such as infection, hypoxia, and inflammation).
Collectively, eBEVs have demonstrated remarkable therapeutic versatility across a broad spectrum of diseases, including intestinal disorders, metabolic diseases, autoimmune conditions, cancer, CNS disorders, and skin injuries. A systematic comparison of disease-specific engineering strategies, therapeutic outcomes, and advantages over non-BEV approaches is provided in Table 3. Despite the largely preclinical nature of current evidence, the rapid pace of innovation in synthetic biology and nanomedicine positions eBEVs as promising candidates for next-generation precision postbiotics, with several candidates poised for clinical translation in the coming years.
Therapeutic applications of eBEVs in various diseases
| Types of diseases | Typical engineering strategies | Key therapeutic outcomes | Stage of development | Comparison with other approaches |
| Intestinal diseases | Genetic engineering for yield enhancement; Surface display of antimicrobial peptide; Physical loading (ultrasound/extrusion) | Restoration of intestinal barrier, modulation of gut microbiota, alleviation of inflammation | Preclinical | Better targeting than conventional anti-inflammatory drugs; Higher safety than live probiotics |
| Metabolic disorders | Genetic engineering for self-assembly loading of therapeutic proteins; Microfluidic sonication loading | Reduction of uric acid, weight loss, amelioration of fatty liver | Preclinical | Higher resistance to gastric degradation than oral enzymes; Better biocompatibility than synthetic NPs |
| Autoimmunity and inflammation | Genetic engineering for co-expression of BMP-2/VEGF; Hydrogel co-encapsulation with IL-4; Thermosensitive hydrogel loading | Promotion of bone regeneration, alleviation of inflammation, modulation of macrophage polarization | Preclinical | Temporal synergy compared to monotherapy; Higher yield, lower cost than stem cell-derived EVs |
| Cancer | Surface display of targeting ligands; Loading of chemotherapeutics/nucleic acids; In situ “living factory” production | Anti-tumor immune activation, tumor regression, inhibition of metastasis | Preclinical | Endogenous adjuvanticity; Higher safety than viral vectors; Improved targeting than conventional chemotherapy |
| CNS disorders | Conceptual stage: BBB-crossing peptides (Angiopep-2); Detoxification (LPS modification); Loading of neurotrophic factors or siRNA | Still in proof-of-concept stage (BBB penetration and neuroprotective potential demonstrated) | Preclinical (concept) | Superior BBB penetration than conventional small molecules and nanocarriers |
| Skin regeneration | Surface display of SOD; Loading of tRNA; Fusion with photothermal hydrogel; Microfluidic artificial vesicles | Promotion of re-epithelialization, angiogenesis, antibacterial effects, anti-photoaging | Preclinical | Higher stability than growth factors; Superior to stem cell therapies (non-animal derived |
CHALLENGES AND FUTURE PERSPECTIVES
Despite demonstrating significant potential in tackling different diseases, eBEVs still face a series of critical challenges in their transition from laboratory to clinical application[181]. Systematically understanding and overcoming these obstacles is essential to achieve their full therapeutic value.
Standardized production of eBEVs
There is a crucial bottleneck concerning production, characterization, and standardization. Although bacteria possess the advantages of rapid proliferation, achieving efficient, stable, and reproducible large-scale production of BEVs remains the primary difficulty[182]. Current laboratory-scale preparation methods (such as ultracentrifugation) are difficult to scale up, and yield and purity of BEVs often cannot be achieved simultaneously. Moreover, different bacterial strains and cultivation conditions (such as stress states) significantly impact BEVs yield and composition, leading to batch-to-batch variability[183]. BEVs constitute a highly heterogeneous population, exhibiting considerable diversity in particle size, membrane composition, and internal cargo (proteins, nucleic acids, metabolites)[184]. The current lack of rapid, precise, and standardized characterization techniques to comprehensively evaluate their physical properties, biochemical composition, drug loading capacity, and functional activity poses significant challenges in establishing unified quality control standards[185].
Biosafety and immunogenicity of eBEVs
Another crucial challenge for the clinical use of eBEVs is the precise regulation of their biosafety and immunogenicity. BEVs from G-/G+ bacteria harbor PAMPs such as LPS and TA/LTA, respectively, on their surfaces, which act as potent immune stimulants[16]. This trait underpins their use as vaccine adjuvants or immunotherapy activators, it may also trigger uncontrolled systemic inflammatory responses, posing significant safety risks. Consequently, achieving detoxification through genetic engineering or chemical modification while preserving essential immunomodulatory functions represents a core challenge in the engineered design. Specific detoxification strategies have been developed, including lipid A engineering (e.g., knockout of msbB, pagL, pagP, or lpxE/lpxF/futB) and flagellin elimination, which can reduce endotoxicity by 10- to 25-fold while preserving adjuvant activity[186,187]. The balance between immunogenicity (desired for vaccines) and toxicity (undesired for drug carriers) is context-dependent and requires careful dose optimization. In Table 4, we provide a summary of balancing strategies across different application scenarios.
A summary of balancing strategies across different application scenarios
| Application scenario | Desired immunogenicity | Acceptable toxicity level | Recommended engineering strategy |
| Cancer vaccine | High (adjuvant effect needed) | Moderate (local inflammation acceptable) | Partial LPS attenuation (msbB knockout); Surface antigen display |
| Infectious disease vaccine | Moderate to high | Low (systemic safety priority) | Lipid A modification (pagL, pagP); Detoxification + alum co-administration |
| Targeted drug delivery | Low (avoid premature clearance) | Minimal | LPS-free OMVs; stealth coating (PEGylation) |
| Autoimmune disease therapy | Low (avoid immune activation) | Minimal | LPS-free OMVs; hybrid vesicles with inert carriers |
Preclinical safety assessment should encompass in vitro cytokine release assays, in vivo acute/chronic toxicity studies, and pyrogenicity testing. Furthermore, the in vivo fate of eBEVs remains incompletely elucidated. They may undergo rapid clearance by the mononuclear phagocyte system or accumulate in non-target tissues, triggering unintended effects[181]. Mitigation strategies include polyethylene glycol modification (PEGylation), targeting ligand conjugation, and biomimetic membrane hybridization. Lastly, comprehensive assessment of long-term biosafety is required, including whether BEVs derived from engineered bacteria could transfer genetic material such as antibiotic resistance genes at the systemic level[185]. Containment strategies, such as non-antibiotic selection markers, chromosomal integration, and suicide switches, can minimize this risk[188].
Collectively, addressing these multifaceted safety and immunogenicity challenges requires an integrated approach combining rational genetic detoxification, rigorous preclinical assessment, advanced biodistribution tracking, and robust containment strategies. Such efforts will be essential to pave the way for the successful clinical translation of eBEVs.
Future perspectives of eBEVs
Based on the challenges outlined above, several promising research directions are anticipated to accelerate the clinical translation of eBEVs. To overcome the production and standardization bottleneck, future efforts should focus on elucidating the molecular mechanisms of BEV biogenesis in different bacteria (particularly probiotics), which will provide theoretical guidance for rational engineering. High‑throughput screening platforms integrating synthetic biology, automation, and artificial intelligence can accelerate the optimization of hypervesiculating strains. Additionally, developing advanced characterization techniques, such as single‑particle analysis and multi‑omics profiling, will enable real‑time quality monitoring and standardization. Also, in response to the safety and immunogenicity concerns, future research should develop “conditional penetration” engineering strategies, such as stimulus‑responsive eBEVs (e.g., low pH, high ROS, specific enzymes) that activate them only in the disease microenvironment. Biomimetic hybrid vesicles that combine BEVs with carriers (e.g., liposomes or cell membranes) offer another avenue to reduce immunogenicity while preserving targeting capabilities. Furthermore, advanced in vivo imaging techniques should be employed to systematically map the biodistribution and clearance kinetics of eBEVs.
The regulatory classification of eBEVs varies across jurisdictions. In the United States, eBEVs are regulated as biological products of the Public Health Service Act by the Food and Drug Administration (FDA)’s Center for Biologics Evaluation and Research (CBER)[189]. In the European Union, they are likely classified as Advanced Therapy Medicinal Products (ATMPs), subject to centralized approval via EMA’s Committee for Advanced Therapies. For first-in-human trials, an Investigational New Drug (IND) application is required in the US, while a Clinical Trial Authorization (CTA) is needed in the Europe (EU), both demanding rigorous chemistry, manufacturing, and control (CMC) data as well as Good Manufacturing Practice (GMP) compliance[182]. Notably, eBEVs derived from genetically modified bacteria face additional regulatory complexity, including biosafety assessments and environmental risk evaluations under genetically modified organism (GMO) legislation. However, ongoing efforts - such as the proposed European Biotech Act - aim to streamline these requirements for ATMPs containing GMOs that pose minimal risk. To pave the way for regulatory and translational approval, international organizations should work toward establishing standardized quality control frameworks and regulatory guidelines tailored to eBEVs. Early engagement with regulatory agencies is encouraged to align development protocols with evolving expectations. Additionally, combining eBEVs with existing therapies (e.g., checkpoint inhibitors, chemotherapies) may accelerate clinical adoption by leveraging their synergistic effects.
In summary, the successful clinical translation of eBEVs will require an integrated, challenge-driven research agenda.
CONCLUSION
In this review, we provide a state of the art scenario of BEVs utilization as postbiotics and directions for their engineered modification, efficacy, and safety. In recent years, EVs have emerged as key participants in cross-cell communication, regulating human health through the diverse bioactive molecules they carry. BEVs possess advantages such as rapid proliferation, abundant gene editing methods, and mature high-density culture techniques, garnering increasing attention, particularly when applied as postbiotics. This approach effectively circumvents various risks associated with injectable therapies, offering a safer and more effective means of improving human health. eBEVs enable the enhancement of their inherent functions, the conferral of novel capabilities, the mitigation of potential risks, increased yield, and improved production efficiency, all while preserving their naturally beneficial components, which facilitates flexible therapeutic strategies, better meeting the demands for health improvement. Emerging evidence indicates that the inherent properties of the parent bacterial strain, including Gram type, envelope structure, LPS/LTA content, basal yield, and cargo composition, can critically determine the baseline characteristics of eBEVs and should guide strain selection for specific therapeutic applications. Tailoring the engineering strategy to a carefully chosen strain thus represents the foundation for successful translation of eBEV‑based precision postbiotics. Looking forward, eBEVs embody the concept of “precision postbiotics” by integrating four engineering-enabled capabilities: targeted delivery, controlled release, programmable cargo loading, and detoxification. This precision framework transforms BEVs from naturally occurring but poorly controllable messengers into customizable therapeutic platforms.
DECLARATIONS
Acknowledgements
All figures in the manuscript, including the graphical abstract, were originally created and drawn by the authors using Adobe Illustrator.
Authors’ contributions
Writing-original draft, Figures drawing: Chen J, Chen Q
Data curation: Li B, Pan C
Writing-review and editing: Grau R, Yang G, Jin Y, Cheng J, Liu T, Li S, Wang Q, Wang Y, Yang Y
Conceptualization: Yang G, Jin Y, Cheng J
Funding acquisition: Cheng J, Liu T, Jin Y
All authors have read and approved the final version of the manuscript and agree to be accountable for all aspects of the work.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported by the Zhejiang Provincial Natural Science Foundation of China (grant number: LQN25C010005), and the financial support from “Pioneer” and “Leading Goose” R&D Program of Zhejiang Province (grant number: 2025C01100), the National Natural Science Foundation of China (grant number: 42277279).
Conflicts of interest
Grau R is affiliated with Kyojin S.A. and Juventas 4life SL, and Yang Y is affiliated with Rawberry Foods Co. Ltd. These affiliations did not influence the design, interpretation, or presentation of this review. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The other authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
2. Bulut SD, Döndaş HA, Celebioglu HU, Sansano JM, Döndaş NY. Recent insights about probiotics related pharmabiotics in pharmacology: prevention and management of diseases. Probiotics Antimicrob Proteins. 2025;17:2262-92.
3. Ma T, Li Y, Yang N, et al. Efficacy of a postbiotic and its components in promoting colonic transit and alleviating chronic constipation in humans and mice. Cell Rep Med. 2025;6:102093.
4. Salminen S, Collado MC, Endo A, et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol. 2021;18:649-67.
5. You J, Liu W, Huang Y, et al. Akkermansia muciniphila PROBIO ameliorates overweight via gut microbiota modulation:a randomized controlled trial. Food Sci Hum Wellness. 2025.
6. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19:213-28.
7. Dorward DW, Garon CF. DNA is packaged within membrane-derived vesicles of gram-negative but not gram-positive bacteria. Appl Environ Microbiol. 1990;56:1960-2.
8. Jahromi LP, Fuhrmann G. Bacterial extracellular vesicles: understanding biology promotes applications as nanopharmaceuticals. Adv Drug Deliv Rev. 2021;173:125-40.
9. Toyofuku M, Nomura N, Eberl L. Types and origins of bacterial membrane vesicles. Nat Rev Microbiol. 2019;17:13-24.
10. Xie J, Li Q, Nie S. Bacterial extracellular vesicles: an emerging postbiotic. Trends Food Sci Technol. 2024;143:104275.
11. Wang S, Gao J, Wang Z. Outer membrane vesicles for vaccination and targeted drug delivery. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2019;11:e1523.
12. Luo R, Chang Y, Liang H, et al. Interactions between extracellular vesicles and microbiome in human diseases: new therapeutic opportunities. Imeta. 2023;2:e86.
13. Li M, Zhou H, Yang C, et al. Bacterial outer membrane vesicles as a platform for biomedical applications: an update. J Control Release. 2020;323:253-68.
14. Kaparakis-Liaskos M, Ferrero RL. Immune modulation by bacterial outer membrane vesicles. Nat Rev Immunol. 2015;15:375-87.
15. Bertani B, Ruiz N. Function and biogenesis of Lipopolysaccharides. EcoSal Plus. 2018;8:ESP-0001.
16. Xie J, Li Q, Haesebrouck F, Van Hoecke L, Vandenbroucke RE. The tremendous biomedical potential of bacterial extracellular vesicles. Trends Biotechnol. 2022;40:1173-94.
17. Sartorio MG, Pardue EJ, Feldman MF, Haurat MF. Bacterial outer membrane vesicles: from discovery to applications. Annu Rev Microbiol. 2021;75:609-30.
18. Li Q, Chen X, Xie J, Nie S. Engineered bacterial extracellular vesicles: developments, challenges, and opportunities. Engineering. 2025;54:291-307.
19. Li R, Helbig L, Fu J, et al. Expressing cytotoxic compounds in Escherichia coli Nissle 1917 for tumor-targeting therapy. Res Microbiol. 2019;170:74-9.
20. Liu H, Geng Z, Su J. Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy. Extracell Vesicles Circ Nucl Acids. 2022;3:63-86.
21. Lieberman LA. Outer membrane vesicles: a bacterial-derived vaccination system. Front Microbiol. 2022;13:1029146.
22. Ji N, Wang F, Wang M, Zhang W, Liu H, Su J. Engineered bacterial extracellular vesicles for central nervous system diseases. J Control Release. 2023;364:46-60.
23. Zheng K, Feng Y, Li L, Kong F, Gao J, Kong X. Engineered bacterial outer membrane vesicles: a versatile bacteria-based weapon against gastrointestinal tumors. Theranostics. 2024;14:761-87.
24. Liu H, Zhang Q, Wang S, Weng W, Jing Y, Su J. Bacterial extracellular vesicles as bioactive nanocarriers for drug delivery: advances and perspectives. Bioact Mater. 2022;14:169-81.
25. Nie X, Li Q, Chen X, Onyango S, Xie J, Nie S. Bacterial extracellular vesicles: vital contributors to physiology from bacteria to host. Microbiol Res. 2024;284:127733.
26. Peregrino ES, Castañeda-Casimiro J, Vázquez-Flores L, et al. The role of bacterial extracellular vesicles in the immune response to pathogens, and therapeutic opportunities. Int J Mol Sci. 2024;25:6210.
27. Bittel M, Reichert P, Sarfati I, et al. Visualizing transfer of microbial biomolecules by outer membrane vesicles in microbe-host-communication in vivo. J Extracell Vesicles. 2021;10:e12159.
28. Schaack B, Mercier C, Katby M, et al. Rapid biodistribution of fluorescent outer-membrane vesicles from the intestine to distant organs via the blood in mice. Int J Mol Sci. 2024;25:1821.
29. Lee KS, Kim Y, Lee JH, et al. Human probiotic lactobacillus paracasei-derived extracellular vesicles improve tumor necrosis factor-α-induced inflammatory phenotypes in human skin. Cells. 2023;12:2789.
30. Han F, Wang W, Shi M, et al. Outer membrane vesicles from bacteria: role and potential value in the pathogenesis of chronic respiratory diseases. Front Cell Infect Microbiol. 2022;12:1093327.
31. Meers PR, Liu C, Chen R, et al. Vesicular delivery of the antifungal antibiotics of lysobacter enzymogenes C3. Appl Environ Microbiol. 2018;84:e01353-18.
32. Kim JH, Jeun EJ, Hong CP, et al. Extracellular vesicle-derived protein from Bifidobacterium longum alleviates food allergy through mast cell suppression. J Allergy Clin Immunol. 2016;137:507-16.e8.
33. Chelakkot C, Choi Y, Kim DK, et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp Mol Med. 2018;50:e450.
34. Ashrafian F, Shahriary A, Behrouzi A, et al. Akkermansia muciniphila-derived extracellular vesicles as a mucosal delivery vector for amelioration of obesity in mice. Front Microbiol. 2019;10:2155.
35. Díez-Sainz E, Milagro FI, Riezu-Boj JI, Lorente-Cebrián S. Effects of gut microbiota-derived extracellular vesicles on obesity and diabetes and their potential modulation through diet. J Physiol Biochem. 2022;78:485-99.
36. Chen CY, Rao SS, Yue T, et al. Glucocorticoid-induced loss of beneficial gut bacterial extracellular vesicles is associated with the pathogenesis of osteonecrosis. Sci Adv. 2022;8:eabg8335.
37. Kang CS, Ban M, Choi EJ, et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One. 2013;8:e76520.
38. Shen Y, Giardino Torchia ML, Lawson GW, Karp CL, Ashwell JD, Mazmanian SK. Outer membrane vesicles of a human commensal mediate immune regulation and disease protection. Cell Host Microbe. 2012;12:509-20.
39. Ma L, Lyu W, Song Y, et al. Anti-inflammatory effect of Clostridium butyricum-derived extracellular vesicles in ulcerative colitis: impact on host microRNAs expressions and gut microbiome profiles. Mol Nutr Food Res. 2023;67:e2200884.
40. Raftar SKA, Ashrafian F, Abdollahiyan S, et al. The anti-inflammatory effects of Akkermansia muciniphila and its derivates in HFD/CCL4-induced murine model of liver injury. Sci Rep. 2022;12:2453.
41. Gu Z, Li F, Liu Y, et al. Exosome-like nanoparticles from Lactobacillus rhamnosus GG protect against alcohol-associated liver disease through intestinal Aryl hydrocarbon receptor in mice. Hepatol Commun. 2021;5:846-64.
42. Jo CS, Myung CH, Yoon YC, et al. The effect of Lactobacillus plantarum extracellular vesicles from Korean women in their 20s on skin aging. Curr Issues Mol Biol. 2022;44:526-40.
43. Gan Y, Chen X, Yi R, Zhao X. Antioxidative and anti-inflammatory effects of Lactobacillus plantarum ZS62 on alcohol-induced subacute hepatic damage. Oxid Med Cell Longev. 2021;2021:7337988.
44. Wang L, Zhao Z, Zhao L, et al. Lactobacillus plantarum DP189 reduces α-SYN aggravation in MPTP-induced Parkinson’s disease mice via regulating oxidative damage, inflammation, and gut microbiota disorder. J Agric Food Chem. 2022;70:1163-73.
45. Ashrafian F, Behrouzi A, Shahriary A, et al. Comparative study of effect of Akkermansia muciniphila and its extracellular vesicles on toll-like receptors and tight junction. Gastroenterol Hepatol Bed Bench. 2019;12:163-8.
46. Liang X, Dai N, Sheng K, et al. Gut bacterial extracellular vesicles: important players in regulating intestinal microenvironment. Gut Microbes. 2022;14:2134689.
47. Wang X, Lin S, Wang L, et al. Versatility of bacterial outer membrane vesicles in regulating intestinal homeostasis. Sci Adv. 2023;9:eade5079.
48. Rodovalho VR, da Luz BSR, Rabah H, et al. Extracellular vesicles produced by the probiotic Propionibacterium freudenreichii CIRM-BIA 129 mitigate inflammation by modulating the NF-κB pathway. Front Microbiol. 2020;11:1544.
49. Choi J, Kim YK, Han PL. Extracellular vesicles derived from Lactobacillus plantarum increase BDNF expression in cultured hippocampal neurons and produce antidepressant-like effects in mice. Exp Neurobiol. 2019;28:158-71.
50. Yang Z, Gao Z, Yang Z, et al. Lactobacillus plantarum-derived extracellular vesicles protect against ischemic brain injury via the microRNA-101a-3p/c-Fos/TGF-β axis. Pharmacol Res. 2022;182:106332.
51. Kim OY, Park HT, Dinh NTH, et al. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat Commun. 2017;8:626.
52. Liu H, Zhang H, Han Y, Hu Y, Geng Z, Su J. Bacterial extracellular vesicles-based therapeutic strategies for bone and soft tissue tumors therapy. Theranostics. 2022;12:6576-94.
53. Qiao L, Rao Y, Zhu K, Rao X, Zhou R. Engineered remolding and application of bacterial membrane vesicles. Front Microbiol. 2021;12:729369.
54. Balhuizen MD, Veldhuizen EJA, Haagsman HP. Outer membrane vesicle induction and isolation for vaccine development. Front Microbiol. 2021;12:629090.
55. Premjani V, Tilley D, Gruenheid S, Le Moual H, Samis JA. Enterohemorrhagic Escherichia coli OmpT regulates outer membrane vesicle biogenesis. FEMS Microbiol Lett. 2014;355:185-92.
56. Elhenawy W, Bording-Jorgensen M, Valguarnera E, Haurat MF, Wine E, Feldman MF. LPS remodeling triggers formation of outer membrane vesicles in salmonella. mBio. 2016;7:e00940-16.
57. Laman JD, ’t Hart BA, Power C, Dziarski R. Bacterial peptidoglycan as a driver of chronic brain inflammation. Trends Mol Med. 2020;26:670-82.
58. Liu Y, Chen J, Raj K, et al. A universal strategy to promote secretion of G+/G- bacterial extracellular vesicles and its application in host innate immune responses. ACS Synth Biol. 2023;12:319-28.
59. Chen J, Liu M, Chen S, et al. Engineered therapeutic bacteria with high-yield membrane vesicle production inspired by eukaryotic membrane curvature for treating inflammatory bowel disease. ACS Nano. 2025;19:2405-18.
60. Moon DC, Choi CH, Lee JH, et al. Acinetobacter baumannii outer membrane protein A modulates the biogenesis of outer membrane vesicles. J Microbiol. 2012;50:155-60.
61. Schwechheimer C, Kulp A, Kuehn MJ. Modulation of bacterial outer membrane vesicle production by envelope structure and content. BMC Microbiol. 2014;14:324.
62. McBroom AJ, Johnson AP, Vemulapalli S, Kuehn MJ. Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J Bacteriol. 2006;188:5385-92.
63. Gong X, Liu S, Xia B, et al. Oral delivery of therapeutic proteins by engineered bacterial type zero secretion system. Nat Commun. 2025;16:1862.
64. McBroom AJ, Kuehn MJ. Release of outer membrane vesicles by Gram-negative bacteria is a novel envelope stress response. Mol Microbiol. 2007;63:545-58.
65. Roier S, Zingl FG, Cakar F, et al. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat Commun. 2016;7:10515.
66. Zingl FG, Kohl P, Cakar F, et al. Outer membrane vesiculation facilitates surface exchange and in vivo adaptation of Vibrio cholerae. Cell Host Microbe. 2020;27:225-37.e8.
67. Mozaheb N, Mingeot-Leclercq MP. Membrane vesicle production as a bacterial defense against stress. Front Microbiol. 2020;11:600221.
68. Gerritzen MJH, Martens DE, Uittenbogaard JP, Wijffels RH, Stork M. Sulfate depletion triggers overproduction of phospholipids and the release of outer membrane vesicles by Neisseria meningitidis. Sci Rep. 2019;9:4716.
69. van de Waterbeemd B, Zomer G, van den Ijssel J, et al. Cysteine depletion causes oxidative stress and triggers outer membrane vesicle release by Neisseria meningitidis; implications for vaccine development. PLoS One. 2013;8:e54314.
70. Gerritzen MJH, Maas RHW, van den Ijssel J, et al. High dissolved oxygen tension triggers outer membrane vesicle formation by Neisseria meningitidis. Microb Cell Fact. 2018;17:157.
71. Mashburn-Warren L, Howe J, Garidel P, et al. Interaction of quorum signals with outer membrane lipids: insights into prokaryotic membrane vesicle formation. Mol Microbiol. 2008;69:491-502.
72. McMahon KJ, Castelli ME, García Vescovi E, Feldman MF. Biogenesis of outer membrane vesicles in Serratia marcescens is thermoregulated and can be induced by activation of the Rcs phosphorelay system. J Bacteriol. 2012;194:3241-9.
73. Bonnington KE, Kuehn MJ. Outer membrane vesicle production facilitates LPS remodeling and outer membrane maintenance in salmonella during environmental transitions. mBio. 2016;7:e01532-16.
74. Gamalier JP, Silva TP, Zarantonello V, Dias FF, Melo RC. Increased production of outer membrane vesicles by cultured freshwater bacteria in response to ultraviolet radiation. Microbiol Res. 2017;194:38-46.
75. Roberts R, Moreno G, Bottero D, et al. Outer membrane vesicles as acellular vaccine against pertussis. Vaccine. 2008;26:4639-46.
76. Bottero D, Zurita ME, Gaillard ME, Bartel E, Vercellini C, Hozbor D. Membrane vesicles derived from bordetella bronchiseptica: active constituent of a new vaccine against infections caused by this pathogen. Appl Environ Microbiol. 2018;84:e01877-17.
77. Wang S, Gao J, Li M, Wang L, Wang Z. A facile approach for development of a vaccine made of bacterial double-layered membrane vesicles (DMVs). Biomaterials. 2018;187:28-38.
78. van der Pol L, Stork M, van der Ley P. Outer membrane vesicles as platform vaccine technology. Biotechnol J. 2015;10:1689-706.
79. van de Waterbeemd B, Zomer G, Kaaijk P, et al. Improved production process for native outer membrane vesicle vaccine against Neisseria meningitidis. PLoS One. 2013;8:e65157.
80. Thomas KJ 3rd, Rice CV. Revised model of calcium and magnesium binding to the bacterial cell wall. Biometals. 2014;27:1361-70.
81. Clifton LA, Skoda MW, Le Brun AP, et al. Effect of divalent cation removal on the structure of gram-negative bacterial outer membrane models. Langmuir. 2015;31:404-12.
82. Maredia R, Devineni N, Lentz P, et al. Vesiculation from Pseudomonas aeruginosa under SOS. ScientificWorldJournal. 2012;2012:402919.
83. Bauwens A, Kunsmann L, Karch H, Mellmann A, Bielaszewska M. Antibiotic-mediated modulations of outer membrane vesicles in enterohemorrhagic Escherichia coli O104:H4 and O157:H7. Antimicrob Agents Chemother. 2017;61:e00937-17.
84. Yun SH, Park EC, Lee SY, et al. Antibiotic treatment modulates protein components of cytotoxic outer membrane vesicles of multidrug-resistant clinical strain, Acinetobacter baumannii DU202. Clin Proteomics. 2018;15:28.
85. Manning AJ, Kuehn MJ. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011;11:258.
86. Balhuizen MD, Versluis CM, van Harten RM, et al. PMAP-36 reduces the innate immune response induced by Bordetella bronchiseptica-derived outer membrane vesicles. Curr Res Microb Sci. 2021;2:100010.
87. Rivera J, Cordero RJ, Nakouzi AS, Frases S, Nicola A, Casadevall A. Bacillus anthracis produces membrane-derived vesicles containing biologically active toxins. Proc Natl Acad Sci U S A. 2010;107:19002-7.
88. Sun B, Lv Z, Li R, et al. Synergistic dual-targeting bacterial extracellular vesicles delivering paclitaxel for precision glioblastoma therapy. Nano Lett. 2025;25:15468-77.
89. Kesty NC, Kuehn MJ. Incorporation of heterologous outer membrane and periplasmic proteins into Escherichia coli outer membrane vesicles. J Biol Chem. 2004;279:2069-76.
90. Kim SH, Kim KS, Lee SR, et al. Structural modifications of outer membrane vesicles to refine them as vaccine delivery vehicles. Biochim Biophys Acta. 2009;1788:2150-9.
91. Ojima Y, Yamaguchi K, Taya M. Quantitative evaluation of recombinant protein packaged into outer membrane vesicles of Escherichia coli cells. Biotechnol Prog. 2018;34:51-7.
92. Thakur M, Dean SN. Engineering outer membrane vesicles to carry enzymes: encapsulation, isolation, characterization, and modification. Methods Mol Biol. 2024;2843:177-94.
93. Galenkamp NS, Van Meervelt V, Mutter NL, van der Heide NJ, Wloka C, Maglia G. Preparation of cytolysin A (ClyA) nanopores. Methods Mol Biol. 2021;2186:11-8.
94. Fahie M, Romano FB, Chisholm C, Heuck AP, Zbinden M, Chen M. A non-classical assembly pathway of Escherichia coli pore-forming toxin cytolysin A. J Biol Chem. 2013;288:31042-51.
95. Murase K. Cytolysin A (ClyA): a bacterial virulence factor with potential applications in nanopore technology, vaccine development, and tumor therapy. Toxins. 2022;14:78.
96. Li Y, Zhao R, Cheng K, et al. Bacterial outer membrane vesicles presenting programmed death 1 for improved cancer immunotherapy via immune activation and checkpoint inhibition. ACS Nano. 2020;14:16698-711.
97. Tang S, Tang D, Zhou H, et al. Bacterial outer membrane vesicle nanorobot. Proc Natl Acad Sci U S A. 2024;121:e2403460121.
98. Thomas SC, Madaan T, Kamble NS, Siddiqui NA, Pauletti GM, Kotagiri N. Engineered bacteria enhance immunotherapy and targeted therapy through stromal remodeling of tumors. Adv Healthc Mater. 2022;11:e2101487.
99. Daleke-Schermerhorn MH, Felix T, Soprova Z, et al. Decoration of outer membrane vesicles with multiple antigens by using an autotransporter approach. Appl Environ Microbiol. 2014;80:5854-65.
100. Chen J, Wei C, Pan C, et al. Genetically engineered Bacillus subtilis-derived extracellular vesicles Alleviates Ulcerative colitis by restoring intestinal barrier and regulating gut microbiota. Chem Eng J. 2025;512:162727.
101. Valguarnera E, Feldman MF. Glycoengineered outer membrane vesicles as a platform for vaccine development. Methods Enzymol. 2017;597:285-310.
102. Price NL, Goyette-Desjardins G, Nothaft H, et al. Glycoengineered outer membrane vesicles: a novel platform for bacterial vaccines. Sci Rep. 2016;6:24931.
103. Chen L, Valentine JL, Huang CJ, et al. Outer membrane vesicles displaying engineered glycotopes elicit protective antibodies. Proc Natl Acad Sci U S A. 2016;113:E3609-18.
104. Long Q, Zheng P, Zheng X, et al. Engineered bacterial membrane vesicles are promising carriers for vaccine design and tumor immunotherapy. Adv Drug Deliv Rev. 2022;186:114321.
105. Cheng L, Peng D, Liu Z, et al. Engineered bacterial outer membrane vesicles enhanced tumor immunotherapy through remodeling tumor stroma and targeted delivery of CD73 siRNA. Biomaterials. 2026;327:123725.
106. Siadat SD, Vaziri F, Eftekhary M, et al. Preparation and evaluation of a new lipopolysaccharide-based conjugate as a vaccine candidate for brucellosis. Osong Public Health Res Perspect. 2015;6:9-13.
107. Micoli F, Alfini R, Di Benedetto R, et al. GMMA is a versatile platform to design effective multivalent combination vaccines. Vaccines. 2020;8:540.
108. Di Benedetto R, Alfini R, Carducci M, et al. Novel simple conjugation chemistries for decoration of GMMA with heterologous antigens. Int J Mol Sci. 2021;22:10180.
109. van den Berg van Saparoea HB, Houben D, Kuijl C, Luirink J, Jong WSP. Combining protein ligation systems to expand the functionality of semi-synthetic outer membrane vesicle nanoparticles. Front Microbiol. 2020;11:890.
110. Peng LH, Wang MZ, Chu Y, et al. Engineering bacterial outer membrane vesicles as transdermal nanoplatforms for photo-TRAIL-programmed therapy against melanoma. Sci Adv. 2020;6:eaba2735.
111. Zheng L, Hu B, Zhao D, et al. Recent progresses of exosome-liposome fusions in drug delivery. Chin Chem Lett. 2024;35:108647.
112. Nsairat H, Khater D, Sayed U, Odeh F, Al Bawab A, Alshaer W. Liposomes: structure, composition, types, and clinical applications. Heliyon. 2022;8:e09394.
113. Zhai Y, Ma Y, Pang B, et al. A cascade targeting strategy based on modified bacterial vesicles for enhancing cancer immunotherapy. J Nanobiotechnology. 2021;19:434.
114. Pan X, Xian P, Li Y, et al. Chemotaxis-driven hybrid liposomes recover intestinal homeostasis for targeted colitis therapy. J Control Release. 2025;380:829-45.
115. Zhao X, Zhao R, Nie G. Nanocarriers based on bacterial membrane materials for cancer vaccine delivery. Nat Protoc. 2022;17:2240-74.
116. Wang D, Liu C, You S, et al. Bacterial vesicle-cancer cell hybrid membrane-coated nanoparticles for tumor specific immune activation and photothermal therapy. ACS Appl Mater Interfaces. 2020;12:41138-47.
117. Guo R, Du F, Xiang X, et al. Ultrasound-enhanced Pt-coordinated polymer immunopotentiators and heterogenic fusion membrane-based multifunctional tumor vaccine nanoplatforms for melanoma treatment. Signal Transduct Target Ther. 2025;10:278.
118. Wang F, Fan J, Pan W, et al. Probiotic-inspired hybrid nanovesicles for enhancing immune checkpoint therapy efficiency via tumor immune microenvironment modulation. Bioact Mater. 2026;56:197-216.
119. Liu G, Huang W, Chen L, et al. Commensal bacterial hybrid nanovesicles improve immune checkpoint therapy in pancreatic cancer through immune and metabolic reprogramming. Nano Today. 2023;52:101993.
120. Ayed Z, Cuvillier L, Dobhal G, Goreham R. Electroporation of outer membrane vesicles derived from Pseudomonas aeruginosa with gold nanoparticles. SN Appl Sci. 2019;1:1600.
121. Guo Q, Li X, Zhou W, et al. Sequentially triggered bacterial outer membrane vesicles for macrophage metabolism modulation and tumor metastasis suppression. ACS Nano. 2021;15:13826-38.
122. Li Y, Zhang K, Wu Y, et al. Antigen capture and immune modulation by bacterial outer membrane vesicles as in situ vaccine for cancer immunotherapy post-photothermal therapy. Small. 2022;18:e2107461.
123. Liang D, Liu C, Li Y, et al. Engineering fucoxanthin-loaded probiotics’ membrane vesicles for the dietary intervention of colitis. Biomaterials. 2023;297:122107.
124. Ban W, Sun M, Huang H, et al. Engineered bacterial outer membrane vesicles encapsulating oncolytic adenoviruses enhance the efficacy of cancer virotherapy by augmenting tumor cell autophagy. Nat Commun. 2023;14:2933.
125. Huang W, Zhang Q, Li W, et al. Development of novel nanoantibiotics using an outer membrane vesicle-based drug efflux mechanism. J Control Release. 2020;317:1-22.
126. Didiot MC, Hall LM, Coles AH, et al. Exosome-mediated delivery of hydrophobically modified siRNA for huntingtin mRNA silencing. Mol Ther. 2016;24:1836-47.
127. Yuan J, Yang J, Hu Z, et al. Safe staphylococcal platform for the development of multivalent nanoscale vesicles against viral infections. Nano Lett. 2018;18:725-33.
128. Bartolini E, Ianni E, Frigimelica E, et al. Recombinant outer membrane vesicles carrying Chlamydia muridarum HtrA induce antibodies that neutralize chlamydial infection in vitro. J Extracell Vesicles. 2013;2:20181.
129. Fantappiè L, de Santis M, Chiarot E, et al. Antibody-mediated immunity induced by engineered Escherichia coli OMVs carrying heterologous antigens in their lumen. J Extracell Vesicles. 2014;3:24015.
130. Han HS, Hwang S, Choi SY, et al. Roseburia intestinalis-derived extracellular vesicles ameliorate colitis by modulating intestinal barrier, microbiome, and inflammatory responses. J Extracell Vesicles. 2024;13:e12487.
131. Chen Q, Fang Z, Yang Z, et al. Lactobacillus plantarum-derived extracellular vesicles modulate macrophage polarization and gut homeostasis for alleviating ulcerative colitis. J Agric Food Chem. 2024;72:14713-26.
132. Tao S, Song M, Fan J, Zhu F, Lv T, Wei H. Lactobacillus johnsonii-derived extracellular vesicles carrying GAPDH protect against ulcerative colitis through modulating macrophage polarization. J Adv Res. 2026;81:409-24.
133. Pirolli NH, Levy D, Schledwitz A, et al. Genetically-programmed hypervesiculation of Lactiplantibacillus plantarum increases production of bacterial extracellular vesicles with therapeutic efficacy in a preclinical inflammatory bowel disease model. Adv Sci. 2026;13:e12679.
134. Pourtalebi Jahromi L, Kronast B, Munkert J, et al. Bioengineered bacterial vesicles and biomimetic hybrids eliminate biofilms and balance the gut microbiome. Small. 2026;22:e2502551.
135. Liu W, Yang J, Wei Z, et al. miRNA-loaded biomimetic nanoparticles orchestrate gut microbe to ameliorate inflammatory bowel disease. Sci Adv. 2025;11:eadw5984.
136. Wang M, Nian L, Zhang S, et al. Probiotics behavior‐inspired biomimetic platform for ulcerative colitis prevention and therapy. Adv Funct Mater. 2025;35:2505817.
137. Liu Q, Li B, Ma J, et al. Development of a recombinant outer membrane vesicles (OMVs)-based vaccine against Helicobacter pylori infection in mice. J Extracell Vesicles. 2025;14:e70085.
138. Liu Q, Li B, Lu J, et al. Recombinant outer membrane vesicles delivering eukaryotic expression plasmid of cytokines act as enhanced adjuvants against Helicobacter pylori infection in mice. Infect Immun. 2023;91:e0031323.
140. Pearlman BL. How would you manage this patient with nonalcoholic fatty liver disease? Ann Intern Med. 2019;171:861-2.
141. Galicia-Garcia U, Benito-Vicente A, Jebari S, et al. Pathophysiology of type 2 diabetes mellitus. Int J Mol Sci. 2020;21:6275.
142. Liu R, Zhang F, He X, Huang K. Plant derived exosome-like nanoparticles and their therapeutic applications in glucolipid metabolism diseases. J Agric Food Chem. 2025;73:6385-99.
143. Wang L, Liang A, Huang J. Exendin-4-enriched exosomes from hUCMSCs alleviate diabetic nephropathy via gut microbiota and immune modulation. Front Microbiol. 2024;15:1399632.
144. Kim JS, Song BJ, Cho YE. Pomegranate-derived exosome-like nanovesicles containing ellagic acid alleviate gut leakage and liver injury in MASLD. Food Sci Nutr. 2025;13:e70088.
145. Bajaj G, Choudhary D, Singh V, et al. MicroRNAs dependent G-ELNs based intervention improves glucose and fatty acid metabolism while protecting pancreatic β-cells in type 2 diabetic mice. Small. 2025;21:e2409501.
146. Han M, Wang J, Zhang Z, et al. Momordica Charantia L.-derived extracellular vesicles achieve pancreatic-targeted delivery and repair insulin-secretory function through dual mechanisms via lymphatic transport. Chem Eng J. 2025;520:165747.
147. Wang K, Li X, Xu X, et al. AI-driven target screening and microfluidic sonication-assembled oral delivery of fucoxanthin-loaded probiotic vesicles for targeted alleviation of diet-induced obesity. Chem Eng J. 2025;518:164598.
148. Lu T, Liu Y, Huang X, et al. Early-responsive immunoregulation therapy improved microenvironment for bone regeneration via engineered extracellular vesicles. Adv Healthc Mater. 2024;13:e2303681.
149. Liu H, Li R, Yang H, et al. Extracellular vesicles in gut-bone axis: novel insights and therapeutic opportunities for osteoporosis. Small Sci. 2025;5:2400474.
150. Han R, Zhou D, Ji N, et al. Folic acid-modified ginger-derived extracellular vesicles for targeted treatment of rheumatoid arthritis by remodeling immune microenvironment via the PI3K-AKT pathway. J Nanobiotechnology. 2025;23:41.
151. Liu Y, Tao S, Zhang Z, et al. Perilla frutescens leaf-derived extracellular vesicle-like particles carry Pab-miR-396a-5p to alleviate psoriasis by modulating IL-17 signaling. Research. 2025;8:0675.
152. Dell’Atti F, Abreu H, Malfa P, Raineri D, Cappellano G, Chiocchetti A. Probiotic-derived extracellular vesicles: the next breakthrough in postbiotics for rheumatoid arthritis. Front Immunol. 2025;16:1620185.
153. Liu H, Song P, Zhang H, et al. Synthetic biology-based bacterial extracellular vesicles displaying BMP-2 and CXCR4 to ameliorate osteoporosis. J Extracell Vesicles. 2024;13:e12429.
154. Zhou G, Zhou Q, Li R, et al. Synthetically engineered bacterial extracellular vesicles and IL-4-encapsulated hydrogels sequentially promote osteoporotic fracture repair. ACS Nano. 2025;19:16064-83.
155. Su D, Li M, Xie Y, et al. Gut commensal bacteria Parabacteroides goldsteinii-derived outer membrane vesicles suppress skin inflammation in psoriasis. J Control Release. 2025;377:127-45.
156. Luo Z, Cheng X, Feng B, et al. Engineering versatile bacteria-derived outer membrane vesicles: an adaptable platform for advancing cancer immunotherapy. Adv Sci. 2024;11:e2400049.
157. Zhuang WR, Xue WC, Liang C, et al. Bacterial foreignization nanosystem elicits multi-phenotypic T cells for antitumor immunity. Adv Sci. 2025;12:e04155.
158. Sun D, Li Y, Yin X, et al. Utilizing engineered bacteria as “cell factories” in vivo for intracellular RNA-loaded outer membrane vesicles’ self-assembly in tumor treatment. ACS Nano. 2024;18:35296-309.
159. Ahn W, Lee Y, Nam GH, You JB, Lee EJ. Bioengineered extracellular vesicles in emerging cancer vaccine platforms. Small Sci. 2025;5:2500270.
160. You H, Zhang S, Zhang Y, et al. Engineered bacterial outer membrane vesicles-based doxorubicin and CD47-siRNA Co-delivery nanoplatform overcomes immune resistance to potentiate the immunotherapy of glioblastoma. Adv Mater. 2025;37:e2418053.
161. Joe E, Ringman JM. Cognitive symptoms of Alzheimer’s disease: clinical management and prevention. BMJ. 2019;367:l6217.
162. Liu J, Wang T, Dong J, Lu Y. The blood-brain barriers: novel nanocarriers for central nervous system diseases. J Nanobiotechnology. 2025;23:146.
163. Rajendran L, Honsho M, Zahn TR, et al. Alzheimer’s disease beta-amyloid peptides are released in association with exosomes. Proc Natl Acad Sci U S A. 2006;103:11172-7.
164. Xin H, Li Y, Cui Y, Yang JJ, Zhang ZG, Chopp M. Systemic administration of exosomes released from mesenchymal stromal cells promote functional recovery and neurovascular plasticity after stroke in rats. J Cereb Blood Flow Metab. 2013;33:1711-5.
165. Xian P, Hei Y, Wang R, et al. Mesenchymal stem cell-derived exosomes as a nanotherapeutic agent for amelioration of inflammation-induced astrocyte alterations in mice. Theranostics. 2019;9:5956-75.
166. Zhang Y, Lu L, Li Y, Liu H, Zhou W, Zhang L. Response surface methodology optimization of exosome-like nanovesicles extraction from lycium ruthenicum murray and their inhibitory effects on Aβ-induced apoptosis and oxidative stress in HT22 cells. Foods. 2024;13:3328.
167. Yu Y, Xu Z, Xu L, Lu D, Tang Y, Mai H. Plant extracellular vesicles as emerging neuroprotective agents for central nervous system disorders. J Adv Res. 2026;79:879-89.
168. Sun B, Sawant H, Borthakur A, Bihl JC. Emerging therapeutic role of gut microbial extracellular vesicles in neurological disorders. Front Neurosci. 2023;17:1241418.
169. Arenas YM, Pérez-Martinez G, Montoliu C, Llansola M, Felipo V. Extracellular vesicles from L. paracasei improve neuroinflammation, GABA neurotransmission and motor incoordination in hyperammonemic rats. Brain Behav Immun. 2025;123:556-70.
170. Kwon H, Lee EH, Park SY, et al. Lactobacillus-derived extracellular vesicles counteract Aβ42-induced abnormal transcriptional changes through the upregulation of MeCP2 and Sirt1 and improve Aβ pathology in Tg-APP/PS1 mice. Exp Mol Med. 2023;55:2067-82.
171. Qin Y, Chen X, Bao L, et al. Lipid metabolism of apoptotic vesicles accelerates cutaneous wound healing by modulating macrophage function. J Nanobiotechnology. 2025;23:106.
172. Yuan S, Jin M, Zhang Y, et al. Extracellular vesicles in skin: biological function and therapeutic potential. Int J Nanomedicine. 2025;20:11211-33.
173. Wu W, Zhang B, Wang W, et al. Plant-derived exosome-like nanovesicles in chronic wound healing. Int J Nanomedicine. 2024;19:11293-303.
174. Wang J, Li X, Zhao X, et al. Lactobacillus rhamnosus GG-derived extracellular vesicles promote wound healing via miR-21-5p-mediated re-epithelization and angiogenesis. J Nanobiotechnology. 2024;22:644.
175. Li Y, Zheng Z, Kong X, et al. Bacteria extracellular vesicles derived from Lactobacillus reuteri delivering intrinsic miR-21a-5p to accelerate diabetic wound healing. Nano Research. 2025;18:94908083.
176. Chen Y, Huang X, Liu A, et al. Lactobacillus reuteri vesicles regulate mitochondrial function of macrophages to promote mucosal and cutaneous wound healing. Adv Sci. 2024;11:e2309725.
177. Chen J, Li B, Wei C, et al. Bacillus subtilis-derived extracellular vesicles displaying superoxide dismutase exhibit superior antioxidant ability in ameliorating skin damage. Int J Biol Macromol. 2026;339:150031.
178. Oh SY, Kim DY, Lee KY, et al. Streptococcus mutans-derived extracellular vesicles promote skin wound healing via tRNA cargo. J Nanobiotechnology. 2025;23:322.
179. Yuan P, Deng M, Li X, et al. Lactobacillus extracellular vesicle-driven oxygen-releasing photothermal hydrogel reprograms macrophages and promotes angiogenesis to accelerate diabetic wound healing. Bioact Mater. 2025;54:144-58.
180. Kang S, Jeon S, Baek H, et al. Lactobacillus-derived artificial extracellular vesicles for skin rejuvenation and prevention of photo-aging. Biomater Sci. 2025;13:2026-35.
181. Guo J, Huang Z, Wang Q, et al. Opportunities and challenges of bacterial extracellular vesicles in regenerative medicine. J Nanobiotechnology. 2025;23:4.
182. Huang C, Cao W, Zhou S, Deng Y. Biogenesis mechanisms, regulatory strategies, and applications of bacterial extracellular vesicles. Crit Rev Biotechnol. 2025;45:1700-16.
183. Rima M, Dakramanji M, El Hayek E, El Khoury T, Fajloun Z, Rima M. Unveiling the wonders of bacteria-derived extracellular vesicles: From fundamental functions to beneficial applications. Heliyon. 2025;11:e42509.
184. Effah CY, Ding X, Drokow EK, Li X, Tong R, Sun T. Bacteria-derived extracellular vesicles: endogenous roles, therapeutic potentials and their biomimetics for the treatment and prevention of sepsis. Front Immunol. 2024;15:1296061.
185. Marquez-Paradas E, Torrecillas-Lopez M, Barrera-Chamorro L, Del Rio-Vazquez JL, Gonzalez-de la Rosa T, Montserrat-de la Paz S. Microbiota-derived extracellular vesicles: current knowledge, gaps, and challenges in precision nutrition. Front Immunol. 2025;16:1514726.
186. Chen MY, Cheng TW, Pan YC, et al. Endotoxin-free outer membrane vesicles for safe and modular anticancer immunotherapy. ACS Synth Biol. 2025;14:148-60.
187. Bai X, Li C, Qiu J, et al. A “plug-and-display” nanoparticle based on attenuated outer membrane vesicles enhances the immunogenicity of protein antigens. J Control Release. 2025;378:687-700.
188. Wegmann U, Carvalho AL, Stocks M, Carding SR. Use of genetically modified bacteria for drug delivery in humans: revisiting the safety aspect. Sci Rep. 2017;7:2294.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].