


Lipid-associated macrophages in metabolic dysfunction-associated steatotic liver disease: immune regulation of lipid burden
 0
0 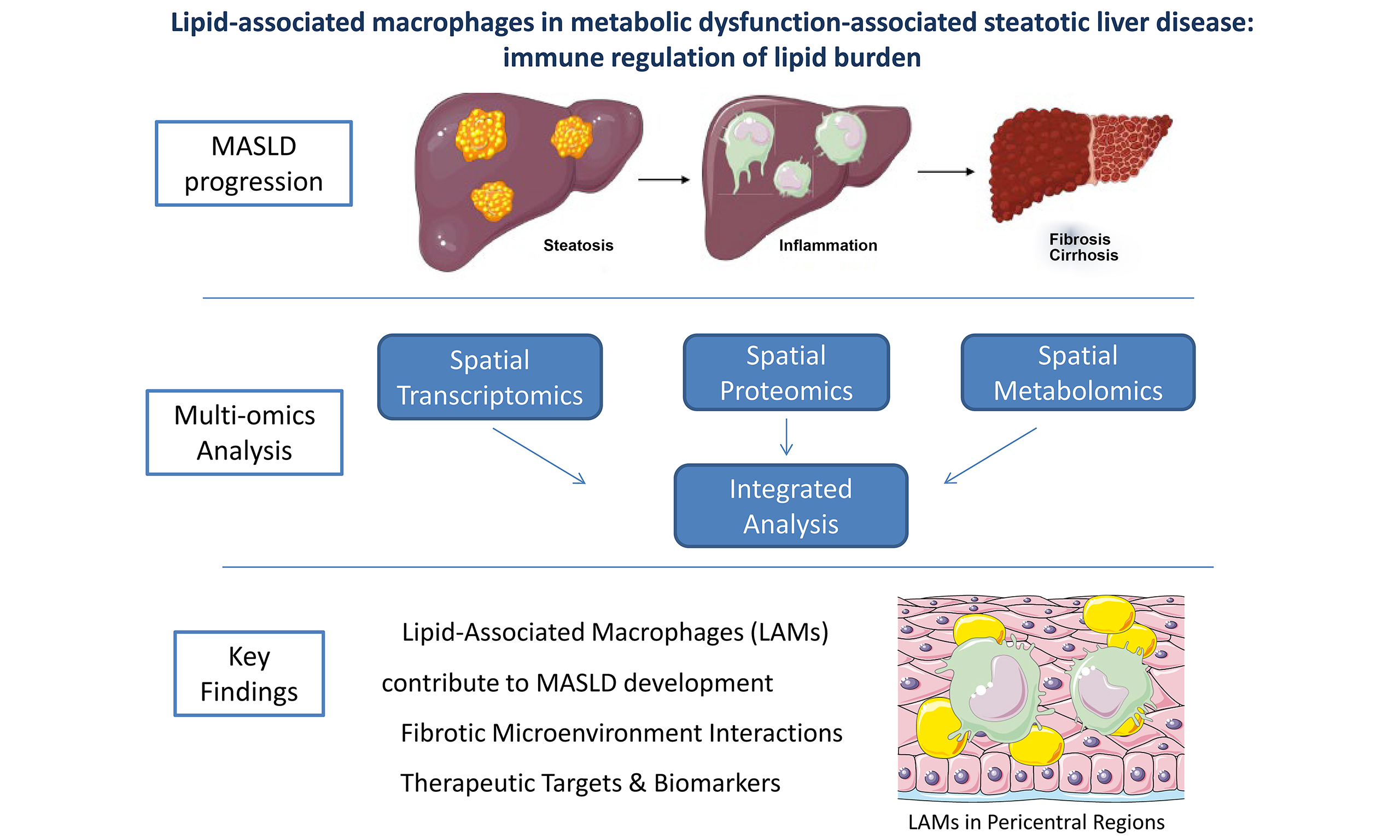
INTRODUCTION
Metabolic dysfunction-associated steatotic liver disease (MASLD), previously known as nonalcoholic fatty liver disease (NAFLD), is a highly prevalent disease[1]. The proposed modifications in terminology, including the adoption of MASLD, metabolic and alcohol-associated liver disease (MetALD) as well as other subtypes under the umbrella of steatotic liver disease (SLD), better captures the underlying complexity of the disease, with important implications for epidemiology, patient classification, and the development of targeted therapies[2]. Today, MASLD is regarded as a systemic disease that impacts multiple organs beyond the liver, including but not limited to the cardiovascular, endocrine, and renal systems; multiple mechanisms, such as insulin resistance, chronic inflammation, and metabolic imbalance, seem to mediate these complex interconnections[3]. MASLD includes a phenotypic spectrum of nosology that can be histologically categorized into: hepatic steatosis, metabolic dysfunction-associated steatohepatitis (MASH), defined as hepatic steatosis and inflammation with hepatocyte injury (hepatocyte ballooning), which may progress to MASH-associated hepatic fibrosis, cirrhosis and hepatocellular carcinoma in a minority of patients[1]. Affecting nearly one in three adults in the general population worldwide, MASLD and its progressive form, MASH, constitute an escalating public health challenge and are projected to become major contributors to advanced liver disease and the need for transplantation[4].
Liver is a multi-functional organ, which is divided into zones. The different zones perform partly distinct tasks through different biological processes[5]. Various structural and immune hepatic cells take part in hepatic functions largely unrelated to those of the hepatocytes[6]. In their study, Li et al. not only investigated molecules that are presented in hepatic tissue of patients with hepatic steatosis or MASH vs. individuals without hepatic disease (controls), but they also focused on their precise spatial localization within hepatic tissue and individual cells[7]. They investigated which genes, proteins and metabolites were expressed and in which specific hepatic regions, rather than assessing only their average levels. To this aim, they applied: (a) single-cell RNA sequencing (scRNA-seq), a technique that isolates whole viable cells from fresh tissue, thus dissociating tissue into individual cells, enabling the characterization of cellular heterogeneity of complex tissue; (b) single-nucleus RNA sequencing (snRNA-seq) analyzes RNA derived from the cell nucleus rather than the whole cell while still enabling reliable identification of cell types; (c) spatial multi-omics, which combines multiple omic layers with their precise location in tissues, thus providing a map of the specific location of molecules[8,9]. More specifically, scRNA-seq offered separate analysis of different cells, such as Kupffer cells, activated hepatic stellate cells (HSCs). Additionally, snRNA-seq offers appropriate preservation of the hepatocytes and lipid cells and more representative cellular composition. The combination of the aforementioned methods seems to be essential, because, as mentioned above, MASLD is characterized by hepatic zonation, heterogeneity of the lesions (steatosis and inflammation predominantly affect the pericentral zone, whereas fibrosis is mainly detected at periportal regions) and the presence of different cells (hepatocytes and multiple immune cells) at different sites[10].
In their study, Li et al. used frozen tissues from liver biopsies obtained from three groups (patient with hepatic steatosis, patients with MASH and apparently healthy controls), which were subjected to spatial transcriptomics, metabolomics and proteomics analyses[7].
THE CORE INSIGHTS OF THE STUDY
Several cells of the hepatic microenvironment are involved in the progression of MASLD[11]. Li et al. created two separate libraries, one for parenchymal cells and one for non-parenchymal cells[7]. A total of 17 different cell types were identified, which were divided into the following categories: (a) hepatic CD45- cells; (b) T-lymphocytes and natural killer (NK) cells; (c) B-lymphocytes and (d) myeloid cells. Furthermore, they divided the hepatic zones into portal, periportal, mid-zonal, and central. It is of interest that Li et al. showed that MASLD modified the dominant types and functions of hepatic immune cells[7]. More specifically, MASLD led to an increase in all T-cell populations, as well as B-cells, and a decrease in the protective resident NK cells. These changes were generally similar in patients with hepatic steatosis and MASH, implying that key immune cell changes or infiltration may occur early in the disease and are maintained when the disease progresses to MASH. However, there were some exceptions. For example, higher numbers of circulating natural killer T (NKΤ) cells were observed in patients with MASLD than controls, thus implying a possible role of NKT cells in the disease progression. Moreover, lipid-associated macrophages (LAMs), a subpopulation of macrophages that accumulate in lipid-rich tissues, such as hepatic steatosis, were more numerous in the liver of patients with MASH than those with hepatic steatosis; LAMs were mainly accumulated in the pericentral region of the liver, thus highlighting the potential contribution of LAMs to the progression of MASLD[7]. It is also of note that high variation in B-cells was observed, implying that the immune response mediated by B-cells was highly individualized among different patients with MASLD.
Following these initial findings, Li et al. tried to investigate the reason why the monocytes infiltrating the liver in MASLD are transformed into LAMs[7]. They showed that microphthalmia-associated transcription factor (MITF), which is highly associated with genes of lipid metabolism, selectively determined the lipid-handling profile and the development of LAMs, being highly activated in MASH patients. Additionally, they observed that peroxisome proliferator-activated receptor-γ (PPAR-γ) signaling pathway was enhanced in LAMs. PPAR-γ acts as a lipid sensor that channels fatty acids towards triglyceride accumulation, improving systemic insulin sensitivity; however, when PPAR-γ is aberrantly activated in the liver, it promotes hepatic steatosis[12,13]. Moreover, spatial transcriptomic analyses revealed that MITF regulates lipid-handling phenotype in LAMs through PPAR-γ driven fatty acid oxidation[7]. Collectively, these findings suggest that MITF, through PPAR-γ, may direct LAMs to the steatotic regions of the liver, thereby contributing to homeostatic adaptation against lipid excess. However, when this process is elongated and/or there is high lipid burden, these macrophages become dysfunctional and may trigger inflammation that may subsequently contribute to the development and progression of fibrosis, instead of tidying lipid excess[14-16]. Therefore, LAMs communicate with other cells and are considered to have initially a hepatoprotective role at the early stages of the disease: by acting as lipid scavengers, they aim to reduce hepatic lipid accumulation. Nevertheless, LAMs may dynamically shift their role from protective tissue repair cells to pro-inflammatory cells, depending on the microenvironment and disease stage[17,18]. Compared with controls, higher inflow and outflow signaling was shown in patients with MASLD by Li et al.[7]. Regarding the comparison between patients with hepatic steatosis and MASH, there were limited differences in incoming signaling; on the contrary, a pronounced expansion of outgoing signaling factors were observed in patients with MASH, including the hepatocyte growth factor (HGF) from LAMs. To frame these alterations, the authors created global intercellular flow networks, not to examine the individual signals, but to map how increased incoming stimuli lead, through specific gene programs, to the production of inflammatory and fibrotic signals by the cells[7]. Additionally, HGF was shown to be potentially hepatoprotective for MASH, by decreasing cell death[7].
Subsequently, the authors proceed with the identification of the specific hepatic regions where fibrosis develops. Spatial validation analysis and proteomics were used to study hepatic tissue from patients with MASLD and controls, and they grouped gene expression into spatial “topics”. They reported that spots with high activity of collagen- or fibrosis-related genes and proteins were localized in histopathologically fibrotic regions and they were connected with HSCs and central vein endothelial cells. These suggest a potential crosstalk between the above cells that may be crucial for fibrogenesis and may be mediated through R-spondin-3 (RSPO3)/leucine-rich repeat-containing G protein-coupled receptor 6 (LGR6) signaling. R-spondins are a group of secreted proteins that potentiate Wnt signaling through interaction with LGRs. Among them, RSPO3 has been shown to play a role in the progression of hepatic fibrosis[19]. In the periportal regions, collagen-related and fibrosis-related gene expression was low at fibrosis stages F1-F2, whereas it was markedly higher at stages F3-F4, thus strengthening the above mentioned speculation.
Based on the aforementioned findings, the authors next integrated spatial analysis with metabolomics to reveal MASLD-specific metabolic programs, particularly in LAMs; more specifically, phospholipid and triglyceride species were strongly upregulated in hepatic pericentral regions of patients with MASLD. MITF-driven expression of phospholipase A2 Group VII (PLA2G7), which was observed exclusively in LAMs, may protect these cells from ferroptosis, thereby possibly sustaining their lipid-handling capacity. It is underlined that PLA2G7 encodes lipoprotein-associated phospholipase A2 (Lp-PLA2), a phospholipid-metabolizing enzyme that is predominantly expressed in macrophages and links lipid metabolism with immune regulation and fibrotic progression in MASLD[20,21]. The above mentioned considerations are depicted in Figure 1.
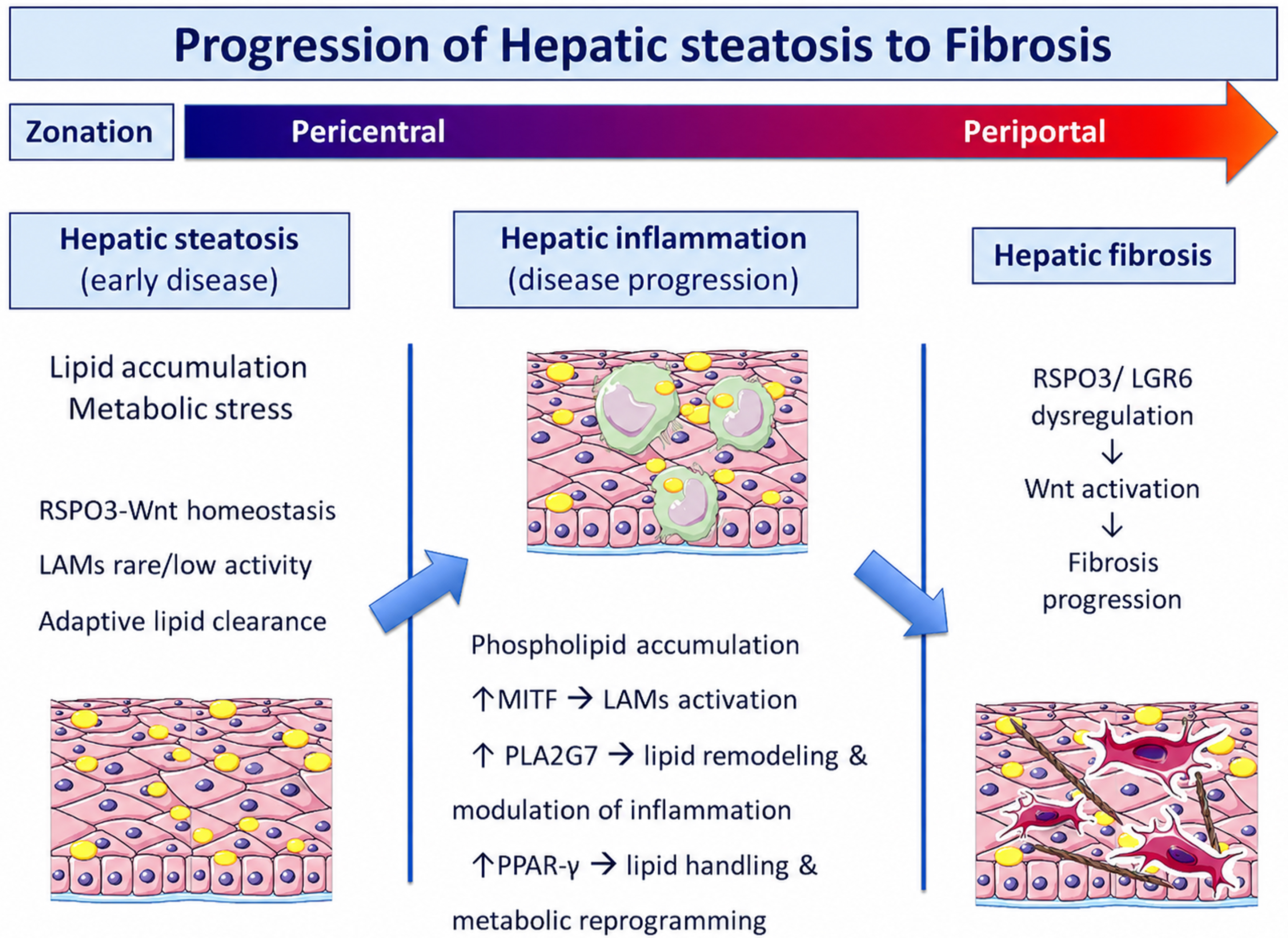
Figure 1. Spatial multi-omics model of MASLD progression from steatosis to fibrosis. In the early phase of MASLD (hepatic steatosis), lipids are accumulated in the hepatocytes. At this phase, inflammatory signaling is minimal and LAMs are present, but they are less abundant and less activated compared with later stages of disease, in which a marked expansion of LAMs is observed. The hepatic microenvironment is characterized primarily by metabolic stress without substantial immune activation. As lipid accumulation intensifies, lipotoxic signals and excess lipid release drive further recruitment and activation of LAMs, particularly within the pericentral (zone 3) region. These macrophages establish a localized inflammatory-metabolic microenvironment, characterized by phospholipid accumulation. LAM activation is associated with upregulation of lipid remodeling and inflammatory pathways, including PLA2G7, as well as transcriptional regulators such as MITF and PPAR-γ. Persistent inflammation and ongoing LAM activity drive stromal remodeling and lead to the development of fibrosis. Crosstalk between endothelial cells and HSCs drives HSC activation and their switch to fibroblasts, leading to extracellular matrix deposition and fibrosis. This process is mediated, in part, by RSPO3-LGR6 dysregulation, which activates Wnt pathways and supports fibrogenic progression. MASLD: Metabolic dysfunction-associated steatotic liver disease; RSPO3: R-spondin 3; Wnt: Wingless/Integrated signaling pathway; LAMs: lipid-associated macrophages; MITF: microphthalmia-associated transcription factor; PLA2G7: phospholipase A2 group VII; PPAR-γ: peroxisome proliferator-activated receptor gamma; LGR6: leucine-rich repeat-containing G protein-coupled receptor 6; HSC: hepatic stellate cell.
DISCUSSION
The involvement of immune cells, such as Kupffer cells and macrophages infiltrating the liver, plays a key role in the progression from hepatic steatosis to MASH[22,23]. Furthermore, inflammation may link MASLD with cardiovascular disease[24]. Macrophages are highly plastic immune cells that may adapt their function and phenotype to environmental changes in the hepatic microenvironment[25]. Although they are traditionally classified into pro-inflammatory (M1) and anti-inflammatory (M2) subtypes, advanced technologies have revealed much greater heterogeneity and additional subtypes[25]. In Li et al. study[7], LAMs were shown to constitute a distinct population of recruited macrophages in the liver of patients with MASLD, especially in MASH as shown in other studies[23]. Although the exact functions of LAMs have not been fully elucidated, it was suggested that they may initially aim to play a protective role by diminishing lipid accumulation[26,27]. The triggering receptor expressed on myeloid cells 2 (TREM2) signaling acts as a key regulator of LAM function[15]. In addition, it is underlined that LAMs may also be referred to as scar-associated or NASH-associated macrophages in other studies[26]. However, under persistently high inflammatory or fibrotic stimuli, sustained overactivation of LAMs has been shown to potentially contribute to activation of inflammatory pathways and enhanced macrophage-mediated inflammation, rather than diminishing hepatic lipid burden[14-16].
Comparing LAMs and Kupffer cells, they seem to express different transcription factors, with MITF possibly being a key regulator of the differentiation of LAMs[7]. Zonated triglyceride accumulation in MASLD is primarily driven by the hepatic oxygen gradient, which limits oxidative metabolism in pericentral regions, thus leading to lipid accumulation rather than oxidation[28,29]. Parenchymal hypoxia, particularly in pericentral regions, may contribute to the preferential accumulation of LAMs in pericentral regions of MASLD[28,29]. Overall, LAMs are predominantly enriched in pericentral regions of the liver, which represent the early sites of lipid accumulation and hepatic inflammation. On the contrary, periportal involvement becomes more pronounced in advanced fibrosis, indicating a spatial expansion of pathological remodeling across the hepatic lobule. These findings may imply a type of coupling among macrophage heterogeneity, hepatic zonation, and disease stage in MASLD[25]. Previous studies have identified LAMs as attractive therapeutic targets in metabolic disease, including MASLD, proposing interventions aiming at regulating their recruitment, functional activation, or lipid-metabolic programs[30].
Concerning fibrosis, Li et al. supported a crosstalk between endothelial cells and HSCs[7], thus connecting inflammation and fibrosis, with RSPO3/LGR6 signaling representing a key regulatory axis and a potential therapeutic target for the prevention or treatment of fibrosis in MASLD[31]. Relevant data support that RSPO3 acts as a significant mediator in the communication among HSCs, hepatocytes, and endothelial cells[32]. This continuous crosstalk aims to secure that the liver maintains its organized structure (zonation) and to counterbalance the effect of various damaging stimuli, i.e., protecting the liver from chronic diseases[31,33]. On the contrary, during chronic injury, the RSPO3 signaling and the subsequent reprogramming of the crosstalk between endothelial cells and HSC seem to favor the fibrogenic cascade, with the HSCs emerging as key collagen producers[32]. In contrast, other studies have reported that downregulation of RSPO3 was associated with impaired liver function and increased fibrosis, highlighting its potential protective role in hepatic homeostasis[33,34]. These apparently conflicting findings underscore the complexity of RSPO3 signaling in liver disease and suggest that further studies are needed to clarify its context-dependent functions. Collectively, LAMs, which are specialized macrophages reflecting metabolic stress, and RSPO3/LGR6 axis, which regulates endothelial-HSC interactions that drive fibrogenesis, may be promising biomarkers for the disease staging and progression, as well as potential therapeutic targets through modulation of LAMs function or restoration of RSPO3/LGR6 signaling.
Notably, Li et al. showed that the region most strongly correlated with MASLD severity, and therefore potentially implicated in the transition from hepatic steatosis to MASH, was enriched in phosphatidylcholine (PC), phosphatidylethanolamine (PE), and phosphatidic acid species, i.e., containing very long-chain fatty acids (C ≥ 22)[7]. Consistent with these human observations, our experimental studies in mice also supported a central role for phospholipid remodeling in hepatic steatosis[35]. More specifically, in a dietary mouse model fed a fast food diet that developed hepatic steatosis, we identified alterations in phospholipids within hepatic tissues. Notably, several phospholipids, including PC, PE, phosphatidylserine, phosphatidylinositol and phosphatidylglycerol species, were different in fast food diet groups compared with the control group, which were fed a chow diet[35]. Importantly, comparable patterns of hepatic phospholipid remodeling have been observed in previous human studies, possibly highlighting that the phospholipid dysregulation may be conserved across species during the progression of MASLD[36-38]. These results demonstrate cross-species consistency, reinforcing the concept that phospholipid remodeling is a relevant feature of MASLD across experimental models and human disease.
One limitation of Li et al.’s study[7] is the unequal sex distribution: the control group consisted only of females, while the MASLD group consisted mainly of males; this may potentially have influenced the results of the study, because of the sexual dimorphism in both hepatic metabolism and immune regulation[39,40]. Data indicate that there are differences in the metabolic profile between men and women, largely attributed to the protective effects of estrogens in premenopausal women[41,42]. Further studies in well balanced groups in terms of sex are warranted to validate the findings of Li et al. Moreover, the small sample size of the study warrants validation of its results in larger populations[7]. Finally, spatial multi-omics approaches are subject to technical limitations, compromising spatial resolution and molecular coverage, as higher resolution often results in fewer detectable molecules and increased data sparsity. In addition, integrating multiple omics layers remains challenging due to differences between data types, technical variability, and computational constraints[43,44].
CONCLUSION
Li et al.’s study provided a comprehensive spatial and single-cell map of MASLD, highlighting the potentially crucial roles of LAMs, of immune cell remodeling, and of phospholipid metabolism in MASH[7]. These findings offer valuable insights into potentially novel biomarkers of disease staging and severity, as well as potential therapeutic targets, such as LAM modulation, for preventing or treating MASLD. Future studies in larger cohorts are warranted to validate and expand these observations.
DECLARATIONS
Authors’ contributions
Conception and design of the study: Makri ES, Polyzos SA
Review of the literature and acquisition of articles: Makri ES
Interpretation of articles: Makri ES, Polyzos SA
Drafting the manuscript: Makri ES, Polyzos SA
Critical revision of the manuscript for important intellectual content: Makri ES, Polyzos SA
Final approval of the version to be submitted: Makri ES, Polyzos SA
All authors have read and agreed to the published version of the manuscript.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
None.
Conflicts of interest
Both authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Tacke F, Horn P, Wai-Sun Wong V, et al. EASL-EASD-EASO Clinical Practice Guidelines on the management of metabolic dysfunction-associated steatotic liver disease (MASLD). Obesity Facts. 2024;17:374-443.
2. Rinella ME, Lazarus JV, Ratziu V, et al.; NAFLD Nomenclature consensus group. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. J Hepatol. 2023;79:1542-56.
3. Targher G, Byrne CD, Lonardo A, Zoppini G, Barbui C. Non-alcoholic fatty liver disease and risk of incident cardiovascular disease: a meta-analysis. J Hepatol. 2016;65:589-600.
4. Chalasani N, Younossi Z, Lavine JE, et al. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology. 2018;67:328-57.
6. Soto-Gutierrez A, Gough A, Vernetti LA, Taylor DL, Monga SP. Pre-clinical and clinical investigations of metabolic zonation in liver diseases: the potential of microphysiology systems. Exp Biol Med. 2017;242:1605-16.
7. Li Z, Luo G, Gan C, et al. Spatially resolved multi-omics of human metabolic dysfunction-associated steatotic liver disease. Nat Genet. 2025;57:3112-25.
8. Vandereyken K, Sifrim A, Thienpont B, Voet T. Methods and applications for single-cell and spatial multi-omics. Nat Rev Genet. 2023;24:494-515.
9. Ding J, Adiconis X, Simmons SK, et al. Systematic comparison of single-cell and single-nucleus RNA-sequencing methods. Nat Biotechnol. 2020;38:737-46.
10. Cunningham RP, Porat-Shliom N. Liver zonation - revisiting old questions with new technologies. Front Physiol. 2021;12:732929.
11. Yang Z, Zhao J, Xie K, Tang C, Gan C, Gao J. MASLD development: from molecular pathogenesis toward therapeutic strategies. Chin Med J. 2025;138:1807-24.
12. Christofides A, Konstantinidou E, Jani C, Boussiotis VA. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism. 2021;114:154338.
13. Grygiel-Górniak B. Peroxisome proliferator-activated receptors and their ligands: nutritional and clinical implications--a review. Nutr J. 2014;13:17.
14. Hu K, Liu S, Sheng S, et al. Heterogeneity of hepatic macrophages in MASLD/MASH: Lipid-associated programs, molecular mechanisms, spatial niches, and therapeutic implications. ILIVER. 2026;5:100234.
15. Shi S, Zhou Y, Zhang H, Zhang J. TREM2 in MASH: integrating lipid metabolism and immune response. Front Immunol. 2025;16:1604837.
16. Xu Y, Hillman H, Chang M, et al. Identification of conserved and tissue-restricted transcriptional profiles for lipid associated macrophages. Commun Biol. 2025;8:953.
17. Govaere O, Petersen SK, Martinez-Lopez N, et al. Macrophage scavenger receptor 1 mediates lipid-induced inflammation in non-alcoholic fatty liver disease. J Hepatol. 2022;76:1001-12.
18. Jaitin DA, Adlung L, Thaiss CA, et al. Lipid-associated macrophages control metabolic homeostasis in a trem2-dependent manner. Cell. 2019;178:686-98.e14.
19. Zhang M, Haughey M, Wang NY, et al. Targeting the Wnt signaling pathway through R-spondin 3 identifies an anti-fibrosis treatment strategy for multiple organs. PLoS One. 2020;15:e0229445.
20. Candels LS, Becker S, Trautwein C. PLA2G7: a new player in shaping energy metabolism and lifespan. Signal Transduct Target Ther. 2022;7:195.
21. Ye J, Song R, Gong X, Li X, Shao C, Zhong B. Fibrosis severity in MASLD determines the predictive value of Lp-PLA2 for carotid atherosclerosis in type 2 diabetes: a cross-sectional study. Biomedicines. 2025;13:2431.
22. Sawada K, Chung H, Softic S, Moreno-Fernandez ME, Divanovic S. The bidirectional immune crosstalk in metabolic dysfunction-associated steatotic liver disease. Cell Metab. 2023;35:1852-71.
23. Ma C, Wang S, Dong B, Tian Y. Metabolic reprogramming of immune cells in MASH. Hepatology. 2025;Epub ahead of print.
24. Lefere S, Tacke F. Macrophages in obesity and non-alcoholic fatty liver disease: Crosstalk with metabolism. JHEP Rep. 2019;1:30-43.
25. Liu W, Li M, Guo H, et al. Single-cell transcriptome analysis of liver immune microenvironment changes induced by microplastics in mice with non-alcoholic fatty liver. Sci Total Environ. 2024;912:168308.
26. De Ponti FF, Liu Z, Scott CL. Understanding the complex macrophage landscape in MASLD. JHEP Rep. 2024;6:101196.
27. Vonderlin J, Chavakis T, Sieweke M, Tacke F. The multifaceted roles of macrophages in NAFLD pathogenesis. Cell Mol Gastroenterol Hepatol. 2023;15:1311-24.
28. Schleicher J, Dahmen U, Guthke R, Schuster S. Zonation of hepatic fat accumulation: insights from mathematical modelling of nutrient gradients and fatty acid uptake. J R Soc Interface. 2017;14:20170443.
29. van der Graaff D, Kwanten WJ, Francque SM. The potential role of vascular alterations and subsequent impaired liver blood flow and hepatic hypoxia in the pathophysiology of non-alcoholic steatohepatitis. Med Hypotheses. 2019;122:188-97.
30. Xu R, Vujić N, Bianco V, et al. Lipid-associated macrophages between aggravation and alleviation of metabolic diseases. Trends Endocrinol Metab. 2024;35:981-95.
31. Chen T, Zhang H, Shan W, Zhou J, You Y. Liver sinusoidal endothelial cells in hepatic fibrosis: opportunities for future strategies. Biochem Biophys Res Commun. 2025;766:151881.
32. Yin X, Yi H, Wang L, Wu W, Wu X, Yu L. RSPOs facilitated HSC activation and promoted hepatic fibrogenesis. Oncotarget. 2016;7:63767-78.
33. Sugimoto A, Saito Y, Wang G, et al. Hepatic stellate cells control liver zonation, size and functions via R-spondin 3. Nature. 2025;640:752-61.
34. Singh A, Akhtar A, Shukla P. Exploring hepatic stellate cell-driven fibrosis: therapeutic advances and future perspectives. ADMET DMPK. 2025;13:2874.
35. Makri ES, Mouskeftara T, Gika H, et al. Serum and liver lipidome following empagliflozin administration for six months in a fast food diet mouse model. Int J Mol Sci. 2025;26:9273.
36. Bertran L, Capellades J, Abelló S, Richart C. Untargeted lipidomic analysis of metabolic dysfunction-associated steatohepatitis in women with morbid obesity. PLoS One. 2025;20:e0318557.
37. Heymann CJF, Mak AL, Holleboom AG, et al. The plasma lipidome varies with the severity of metabolic dysfunction-associated steatotic liver disease. Lipids Health Dis. 2024;23:402.
38. Pebriana RB, Chen T, Derks RJE, et al. Non-invasive identification of steatohepatitis in patients with MASLD using a sterol and lipidomic signature. J Lipid Res. 2025;66:100845.
39. Meyer J, Teixeira AM, Richter S, et al. Sex differences in diet-induced MASLD - are female mice naturally protected? Front Endocrinol. 2025;16:1567573.
40. Makri ES, Xanthopoulos K, Mavrommatis Parasidis P, et al. Partial validation of a six-month high-fat diet and fructose-glucose drink combination as a mouse model of nonalcoholic fatty liver disease. Endocrine. 2024;85:704-16.
41. Ballestri S, Nascimbeni F, Baldelli E, Marrazzo A, Romagnoli D, Lonardo A. NAFLD as a sexual dimorphic disease: role of gender and reproductive status in the development and progression of nonalcoholic fatty liver disease and inherent cardiovascular risk. Adv Ther. 2017;34:1291-326.
42. Cherubini A, Della Torre S, Pelusi S, Valenti L. Sexual dimorphism of metabolic dysfunction-associated steatotic liver disease. Trends Mol Med. 2024;30:1126-36.
43. Geng A, Cui C, Luo Z, et al. Computational methods for spatial multi-omics integration. Biotechnol Adv. 2026;87:108807.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0













Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].