




Immunoregulatory extracellular vesicles in multiple myeloma patients’ peripheral blood
 0
0 
Abstract
Aim: Extracellular vesicles (EVs) mediate cell-to-cell communication and co-contribute in cancer progression and modulation of immune responses. The aim of this work was to characterize the phenotype and function of EVs isolated from the peripheral blood (PB) of multiple myeloma (MM) patients, with particular attention to adenosinergic ectoenzymes.
Methods: EVs were isolated by centrifugation of PB plasma samples from newly diagnosed (ND) MM patients and healthy donors (HD). EVs were characterized by nanoparticle tracking analysis (NTA). Flow cytometry was used to assess the expression of CD138 (a tumor-associated antigen), nicotinamide phosphoribosyltransferase/pre-B cell colony enhancing factor (NAMPT/PBEF1), and adenosinergic ectoenzymes CD38, CD39, CD203a, and CD73. Functional studies were performed by assessing T-cell proliferation in vitro using carboxyfluorescein succinimidyl ester (CFSE) dilution and flow cytometry, with or without EVs to the culture.
Results: The PB EVs size was comparable between MM patients and HDs, whereas EV concentration was significantly higher in MM patients. EVs from MM patients expressed higher levels of CD138, PBEF, CD38, and CD73 than those from HDs. In addition, CD138+ EVs from MM patients expressed higher levels of PBEF, CD38, CD39, and CD73 than CD138- counterparts. Finally, MM-derived EVs inhibited T-cell proliferation in vitro. This inhibition correlated with the expression of CD138 and CD203a.
Conclusion: This study provides the first phenotypic and functional characterization of EVs obtained from the PB of MM patients. The presence of functional adenosinergic molecules may represent a novel immune-escape mechanism operating at distant sites. Nonetheless, further studies are required to define specific contributions of adenosinergic ectoenzymes and other immune-regulatory mechanisms.
Keywords
INTRODUCTION
The clonal expansion of plasma cells (PCs) in the bone marrow (BM) is the hallmark of multiple myeloma (MM), a hematological tumor that typically affects subjects over 60 years of age and represents about 0.9% of all cancers worldwide, contributing to about 1% of cancer-related deaths[1]. Although the etiology of MM has not been fully elucidated, multiple cytogenetic alterations, such as chromosomal deletions and/or translocations, are present and correlated with disease progression and high-risk MM[2]. Asymptomatic forms like monoclonal gammopathy of undetermined significance (MGUS) and smoldering multiple myeloma (SMM) represent premalignant stages that may advance to symptomatic MM. Disease progression is characterized by severe and irreversible impairment of vital organs, including hypercalcemia, renal failure, anemia, and lesions in the bones[2].
The expression and function of adenosinergic pathways in MM have been previously described by our group and others[3,4]. The canonical pathway for adenosine (ADO) production is initiated by the nucleoside triphosphate diphosphohydrolase/CD39, which converts adenosine triphosphate (ATP) into adenosine monophosphate (AMP)[5]. Subsequently, the hydrolysis of AMP to ADO is mediated by ecto-5’-nucleotidase/CD73[6,7]. In contrast, the non-canonical pathway, whose efficiency increases at low pH, starts with the conversion of NAD+ to adenosine diphosphate ribose (ADPR) mediated by the nicotinamide adenine dinucleotide (NAD+)-glycohydrolase/CD38. ADPR is subsequently converted to AMP by CD203a/ENPP1 (PC-1), which has been characterized in both tumor and immune cells[8], although the conversion of ADPR to AMP was first demonstrated functionally by our group[9]. AMP is finally metabolized to ADO by CD73[7,10,11], which catalyzes the final step of both adenosinergic pathways. Functionally, adenosinergic ectoenzymes are key players in the establishment of an immune-suppressive BM microenvironment through a local production of ADO[4,12]. These processes are facilitated by nicotinamide phosphoribosyltransferase/pre-B cell colony enhancing factor (NAMPT/PBEF1), a key enzyme involved in the conversion of nicotinamide and 5-phosphoribosyl-1-pyrophosphate into nicotinamide mononucleotide, which is subsequently transformed into NAD+. This pathway supports not only glycolysis but also the activity of NAD+-dependent enzymes and DNA repair mechanisms[13-16]. NAMPT/PBEF1 expression is increased in both solid and hematological tumors[17,18] as well as in inflammatory diseases[19,20].
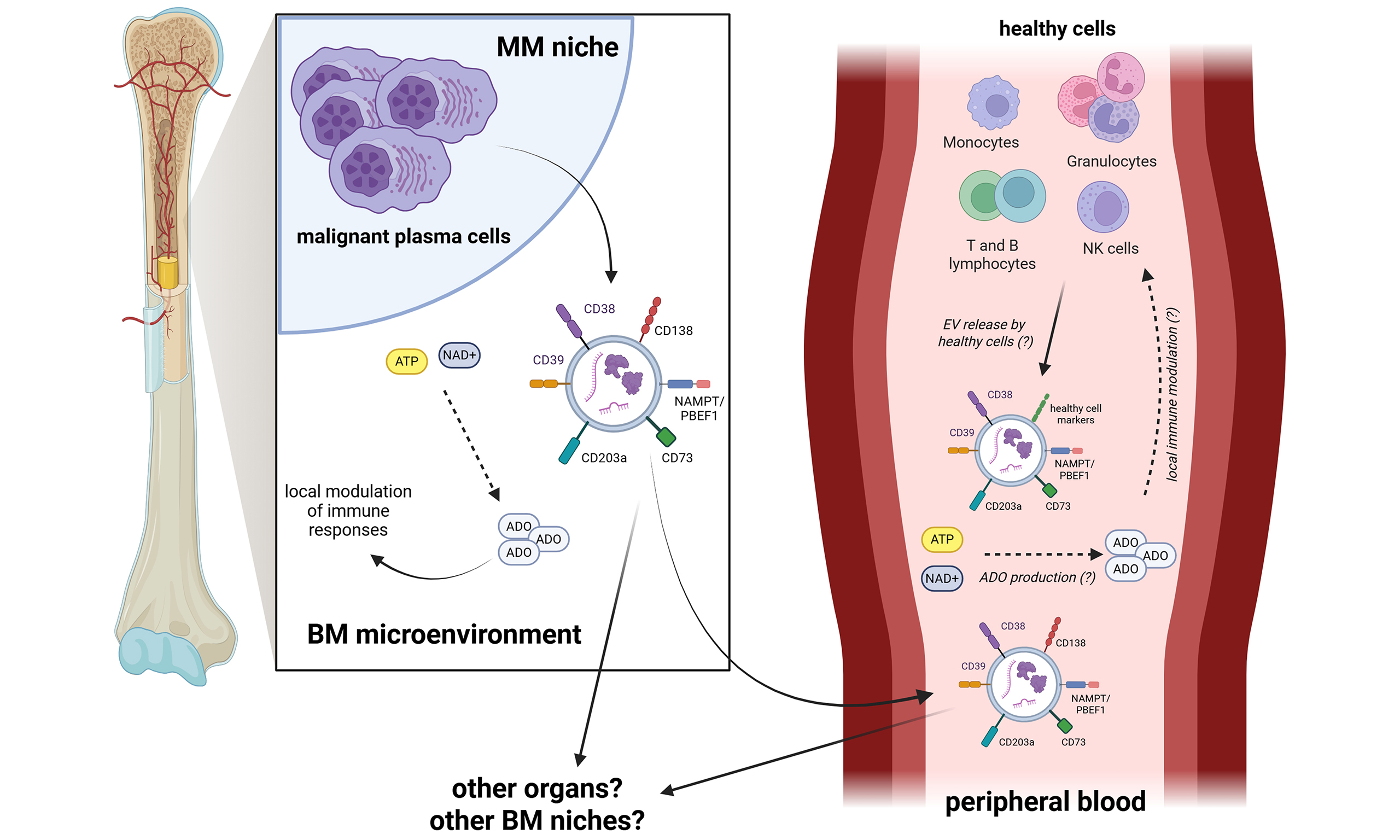
We have recently reported that many of these molecules are also expressed on extracellular vesicles (EVs) obtained from BM plasma samples of MM patients[21]. Several studies have described these EVs as resembling the phenotypic and functional characteristics of their cells of origin. Furthermore, their role in multiple processes related to MM pathogenesis, including modulation of immune responses, progression of the disease, therapy resistance, osteolysis, angiogenesis, and formation of the metastatic niche has been addressed[22-25]. However, to date, no information is available on tumor-derived EVs present in the peripheral blood (PB) of MM patients. Therefore, the aim of this study was to characterize these EVs in terms of size, number, and phenotype, and to assess whether they exert immune-regulatory functions in vitro. The data obtained in this study indicate that tumor-derived EVs can be also detected in the PB of newly diagnosed MM (NDMM) patients and may contribute to tumor-associated immune suppression.
METHODS
Patient characteristics and enrollment criteria
PB samples were collected from 13 NDMM patients enrolled at Istituto di Ricerca e Cura a Carattere Scientifico (IRCCS) Azienda Ospedaliera Metropolitana (AOM), Genova, Italy. Patients were eligible for enrollment if they met the following inclusion criteria: age
Healthy donors’ (HD) PB samples (n = 12) were used as controls. All samples were collected after written informed consent was obtained from all subjects, in accordance with the Declaration of Helsinki. The study was approved by the Ethics Committee (CER Liguria: 409/2021 - DB ID 11602). The clinical features of MM patients are reported in Table 1.
Clinical characteristics of MM patients
| Age at diagnosis | Myeloma subtype | R-ISS-stage | CA |
| 71 | IgG lambda | II | None |
| 77 | IgA kappa | II | del(13q) |
| 71 | IgG kappa | II | del(13q) |
| 81 | IgA kappa | II | N/A |
| 78 | IgA kappa | II | del(13q), (1q) gain |
| 66 | IgG kappa | III | 5p15-9q22-15CEN hyperdiploid status |
| 87 | IgG kappa | II | N/A |
| 82 | IgG lambda | I-II | N/A |
| 62 | IgG kappa | II | N/A |
| 60 | IgA lambda | II | (1q) gain |
| 69 | IgA lambda + IgG kappa | III | N/A |
| 68 | IgG kappa | N/A | N/A |
| 80 | IgG kappa | I | None |
Isolation and characterization of EVs
Blood samples were centrifuged at 3,000 × g for 15 min at 4 °C to obtain PB plasma, which was divided into aliquots (300 μL each) and cryopreserved at -80 °C. Isolation of EVs from thawed samples was performed as previously reported[4]. For some patients, multiple EV preparations were obtained from different plasma aliquots, resulting in a total of 22 EV preparations analyzed. Briefly, PB plasma samples were diluted 1:3 with phosphate buffered saline (PBS), and platelets and debris were removed by centrifugation at 3,000 × g for 15 min at 4 °C. The collected supernatants were analyzed to determine EV size and concentration with the employment of a NanoSight NS500 instrument [nanoparticle tracking analysis (NTA) 2.3 software, 488 nm laser]. Finally, EVs were enriched by centrifugation (20,000 × g at 4 °C for 1 h). After washing with PBS, EVs were resuspended in Magnetic-Activated Cell Sorting (MACS) buffer (PBS/EDTA with 0.5% BSA, 50 μL). For functional experiments, enriched EVs were resuspended in sterile serum-free RPMI-1640 medium.
Flow cytometric analysis
The number of EVs enriched from 300 μL of PB plasma samples, and then subjected to flow cytometric analysis, varied between subjects. CD138 and NAMPT/PBEF1 expression was evaluated using rat Alexa Fluor 488-conjugated anti-CD138 mAb (#359103) and rat Alexa Fluor 647-conjugated anti-PBEF1 mAb (#362616), respectively. The expression of ectoenzymes was evaluated using anti-CD38 (#IB4) and anti-CD73 (#CB73) monoclonal antibodies (mAbs), generated in our laboratory and conjugated with fluorescein isothiocyanate (FITC), phycoerythrin (PE), or allophycocyanin (APC) fluorochromes by Aczon (Bologna, Italy). Anti-CD203a (PC-1, clone #3E8) was kindly provided by J. Goding, and anti-CD39 PE-Cy7 mAbs were purchased from eBiosciences. Irrelevant isotype- and fluorochrome-matched mAbs were obtained from Beckman Coulter.
EVs were incubated with mAbs (20 min at 4 °C in the dark), washed, centrifuged (20,000 × g at 4 °C for 1 h), and resuspended in MACS buffer (200 μL). A Gallios flow cytometer (Beckman Coulter) was used for data acquisition, and Kaluza software version 2.1 (Beckman Coulter) was used for data analysis. At least 2,000 events per sample were acquired using a forward- and side-scatter-based gating strategy to exclude debris background. The expression of ectoenzymes and NAMPT/PBEF1 in EVs specifically derived from MM cells was investigated by gating on CD138+ EVs. Results were expressed as mean relative fluorescence intensity (MRFI), calculated as the ratio between the mean fluorescence intensity obtained with the specific antibody and that obtained with the corresponding irrelevant isotype control.
T-cell proliferation assay
Mononuclear cells (MNCs) were isolated from buffy coat preparations obtained from healthy donors (HDs; n = 3) enrolled at AOM, following written informed consent. Isolation was performed using Ficoll-Paque density gradient centrifugation (800 × g for 30 min at room temperature). The CellTraceTM carboxyfluorescein succinimidyl ester (CFSE) Cell Proliferation Kit (Thermo Fisher Scientific) was used to assess T-cell proliferation. Briefly, MNCs were stained with CFSE
Statistical analysis
Statistical analyses were carried out using GraphPad Prism version 5.03. Data normality was assessed by D’Agostino-Pearson omnibus test. Accordingly, parametric (t-test) or non-parametric (Mann-Whitney or Wilcoxon) tests have been used to compare data. Correlations were investigated by linear regression analysis and Spearman’s rank correlation coefficient. Statistical significance was reached with P values < 0.05.
RESULTS
Characterization of EVs from MM patients and HDs
Sixteen PB plasma samples were obtained from 13 MM patients (one PB sample from 10 patients and two samples from remaining 3 MM patients), as well as from 12 HDs. Plasma samples were characterized for EV size distribution and concentration using NTA. The mean size of EVs was similar between MM patients [black circles; mean ± standard error (SE): 104.8 ± 5.6 nm] and HDs (white circles; 103.8 ± 6.9 nm) [Figure 1A]. In contrast, EVs concentration was significantly higher in plasma samples from MM patients [mean ± standard deviation (SD): 4.1 ± 0.56 × 1011 particles/mL] than in those from HDs (2.0 ± 0.38 × 1011 particles/mL; P = 0.0072) [Figure 1B].
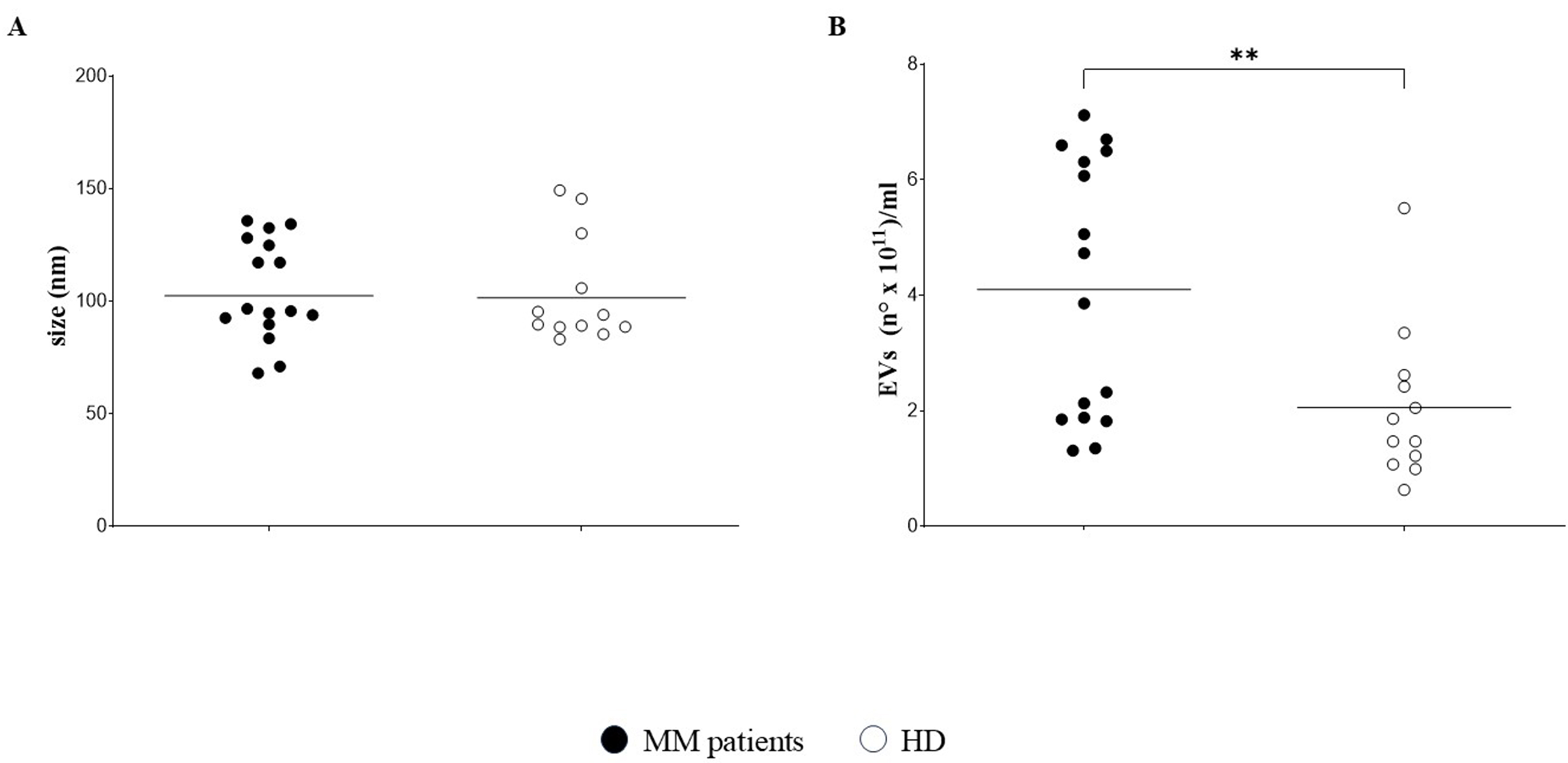
Figure 1. Characterization of EVs. EVs were characterized in 16 PB plasma samples from 13 MM patients and 12 HDs by NTA. Size distribution (nm) is shown in (A), whereas concentration (particles × 1011/mL) is shown in (B). Horizontal bars indicate the means. Asterisk indicates statistically significant differences as calculated by the Mann-Whitney test (**P ≤ 0.01). EVs: Extracellular vesicles; PB: peripheral blood; MM: multiple myeloma; HDs: healthy donors; NTA: nanoparticle tracking analysis.
Expression of CD138, PBEF, CD38 and CD73 is increased in MM patients’ EVs
We next analyzed EVs for surface expression of the tumor-associated marker CD138, as well as NAMPT/PBEF1 and adenosinergic molecules CD38, CD39, CD203a, and CD73. The expression level of CD138, measured as MRFI, was higher in EVs isolated from MM patients (n = 10; MRFI mean ± SE: 7 ± 2.18) than in those from HDs (n = 12; 2.53 ± 0.47; P = 0.04) [Figure 2A]. Similarly, the expression of PBEF1, CD38, and CD73 was significantly higher in EVs from MM patients (14.5 ± 4.8, 484.2 ± 256, and 7.43 ± 2.33, respectively) than in those from HDs (4.7 ± 0.67, P = 0.04; 79 ± 28.8, P = 0.05; and 2.36 ± 0.29, P = 0.01, respectively). In contrast, the expression of CD203a and CD39 was comparable in EVs from MM patients (53.2 ± 28.8 and 36.8 ± 12.3, respectively) and HDs (19.0 ± 15 and 16.5 ± 3.3, respectively). A representative staining is shown in Figure 2B.
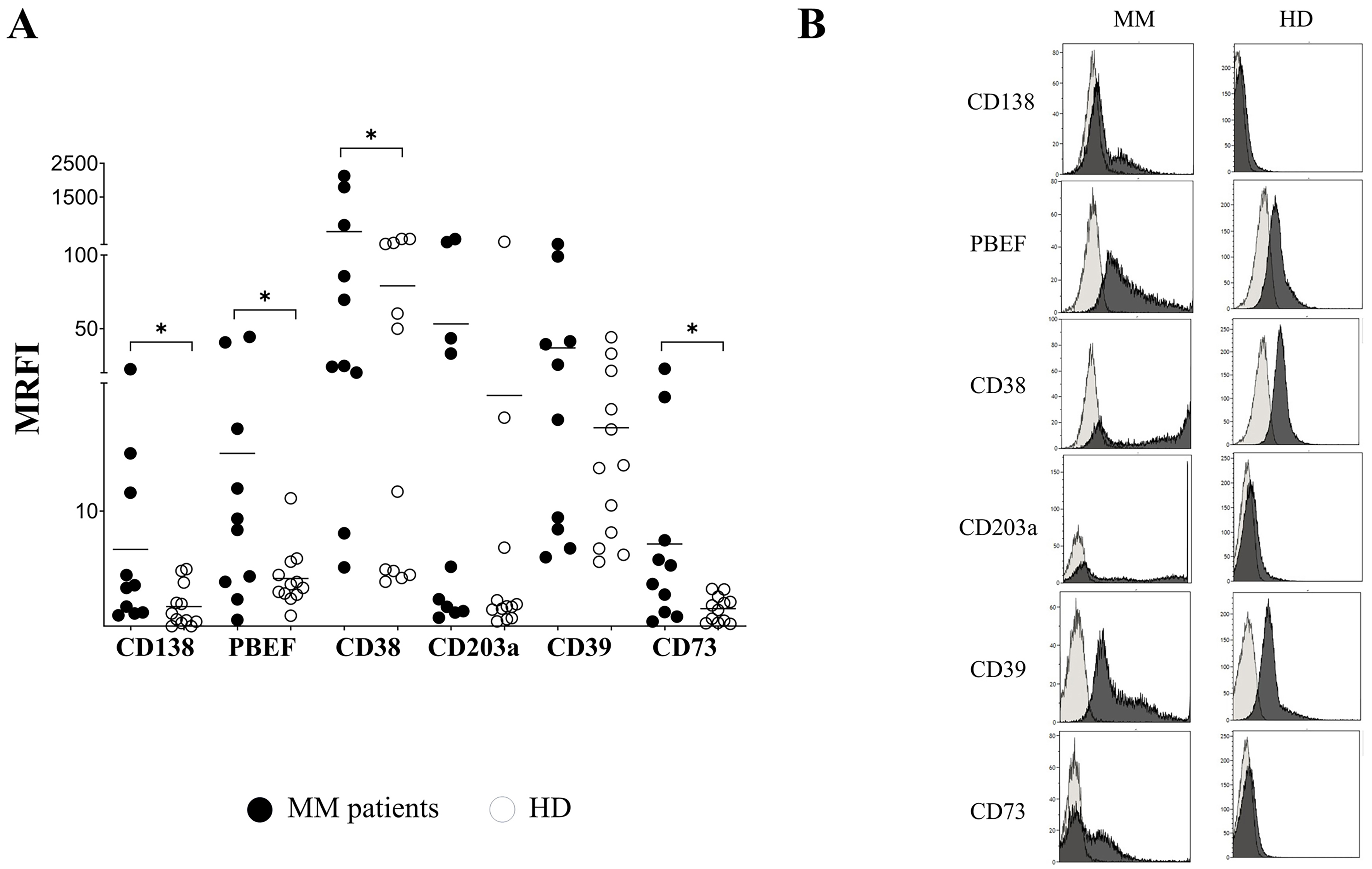
Figure 2. Immunophenotype of EVs. (A) EVs from PB plasma samples obtained from MM patients (n = 10, black dots) and HDs (n = 12, white dots) were analyzed by flow cytometry to assess the expression of CD138, NAMPT/PBEF1 (PBEF), CD38, CD203a, CD39, and CD73. Data are expressed as MRFI. Horizontal bars indicate the mean. Asterisk indicated statistically significant differences calculated using the Mann-Whitney test or unpaired t test, depending on data distribution (*P ≤ 0.05). A representative experiment is shown in (B). Dark grey profiles indicate staining with specific mAbs, whereas light grey profiles indicate staining with irrelevant isotype-matched mAbs. EVs: Extracellular vesicles; PB: peripheral blood; MM: multiple myeloma; HDs: healthy donors; NAMPT/PBEF1 (PBEF): nicotinamide phosphoribosyltransferase/pre-B-cell colony-enhancing factor 1; MRFI: mean relative fluorescence intensity; mAbs: monoclonal antibodies.
Next, we investigated whether phenotypic differences could be observed in EVs derived from malignant PC (identified as CD138+) and those released by healthy cells (i.e., CD138-) in the PB of MM patients. To address this issue, we compared the expression of NAMPT/PBEF1 and adenosinergic ectoenzymes in CD138+ and CD138- EVs across 22 EV preparations from plasma samples of10 MM patients. As shown in Figure 3A, the expression of NAMPT/PBEF1, CD38, CD39, and CD73 was significantly higher in CD138+ (6.2 ± 1.4, 4.89 ± 0.9, 4.0 ± 4.3, and 2.64 ± 0.21, respectively) than in CD138- EVs (1.47 ± 0.24, P < 0.0001; 2.63 ± 0.45, P < 0.001; 1.81 ± 0.39, P = 0.0007; and 1.3 ± 1.34, P = 0.03, respectively). In contrast, the expression of CD203a did not differ significantly between CD138+ (2.48 ± 0.22) and CD138- EVs (1.47 ± 0.39). Taken together, these data suggest that EVs released from malignant MM cells are enriched in molecules with immune-inhibitory potential, an aspect examined in the subsequent section. A representative staining is shown in Figure 3B.
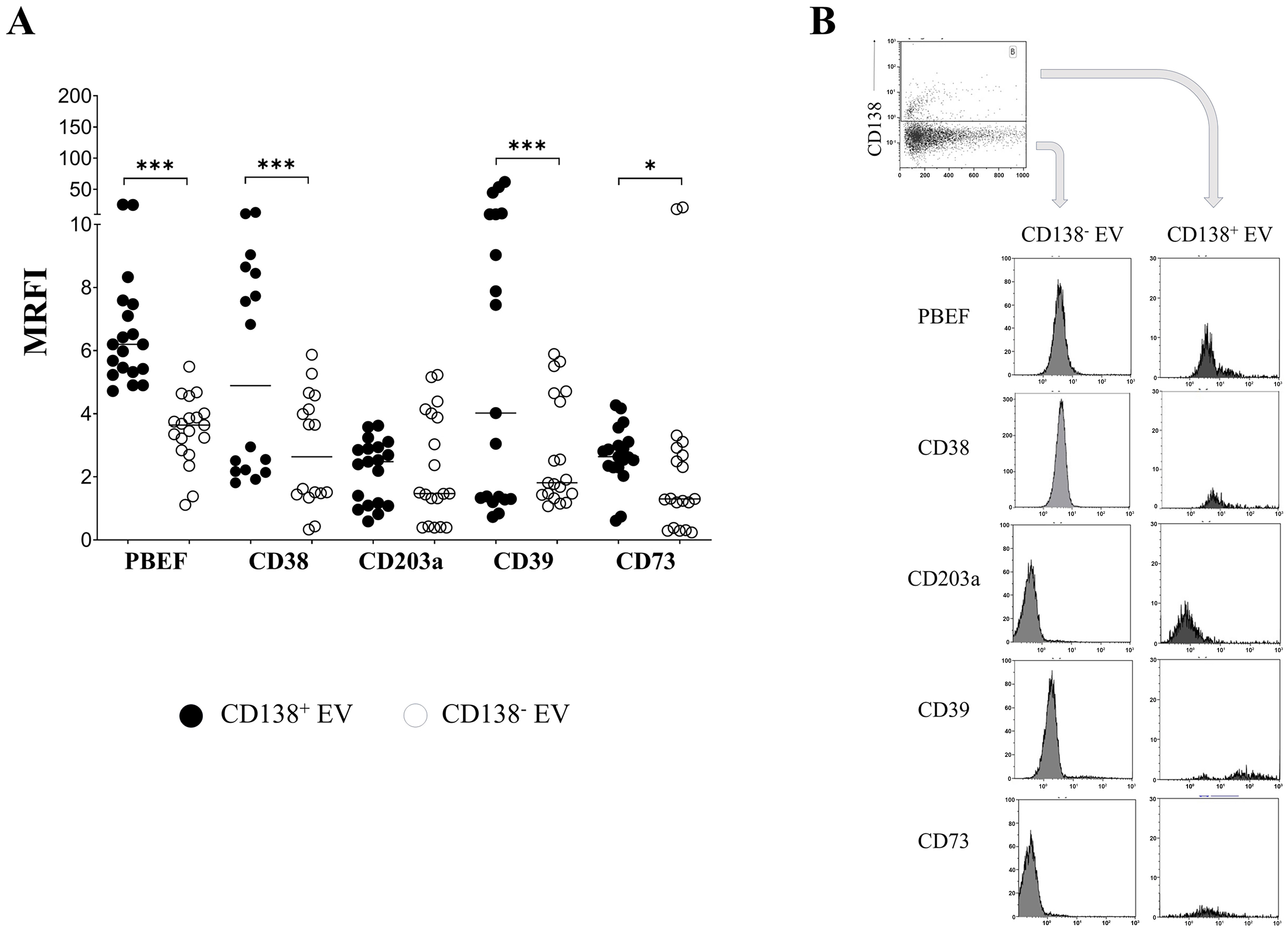
Figure 3. Immunophenotype of CD138+ and CD138- EVs. (A) EVs from PB plasma samples (n = 22) obtained from 13 MM patients were analyzed by flow cytometry to assess the expression of NAMPT/PBEF1 (PBEF), CD38, CD203a, CD39, and CD73 in CD138+ (black dots) and CD138- (white dots) EVs. Data are expressed as MRFI. Horizontal bars indicate the mean. The asterisk indicates statistically significant differences calculated using the Wilcoxon matched-pairs signed-rank test or paired t test, depending on data distribution (*P ≤ 0.05, ***P ≤ 0.001); (B) shows a representative experiment. Dark grey profiles indicate staining with specific mAbs, whereas light grey profiles correspond to staining with irrelevant isotype-matched mAbs. A plot showing CD138 expression on EVs and the gating strategy is shown in the upper panel. EVs: Extracellular vesicles; PB: peripheral blood; MM: multiple myeloma; NAMPT/PBEF1 (PBEF): nicotinamide phosphoribosyltransferase/pre-B-cell colony-enhancing factor 1; MRFI: mean relative fluorescence intensity; mAbs: monoclonal antibodies; CD: cluster of differentiation.
Immune responses are modulated by MM patients’ EVs in vitro
We next tested the ability of EVs obtained from MM patients’ PB plasma samples to regulate immune effector cell functions in vitro. To address this issue, MNCs from HD were stimulated with SEB in the presence or absence of MM-derived EVs. As shown in Figure 4A, CD4⁺ and CD8+ T-cell proliferation after stimulation with SEB (% proliferating cells; mean ± SD: 86.5 ± 4.0 and 79.3 ± 10.8, respectively) was downregulated when MM-derived EVs were added (61.2 ± 16.3, P = 0.01 and 54.8 ± 15.5, P = 0.01, respectively).
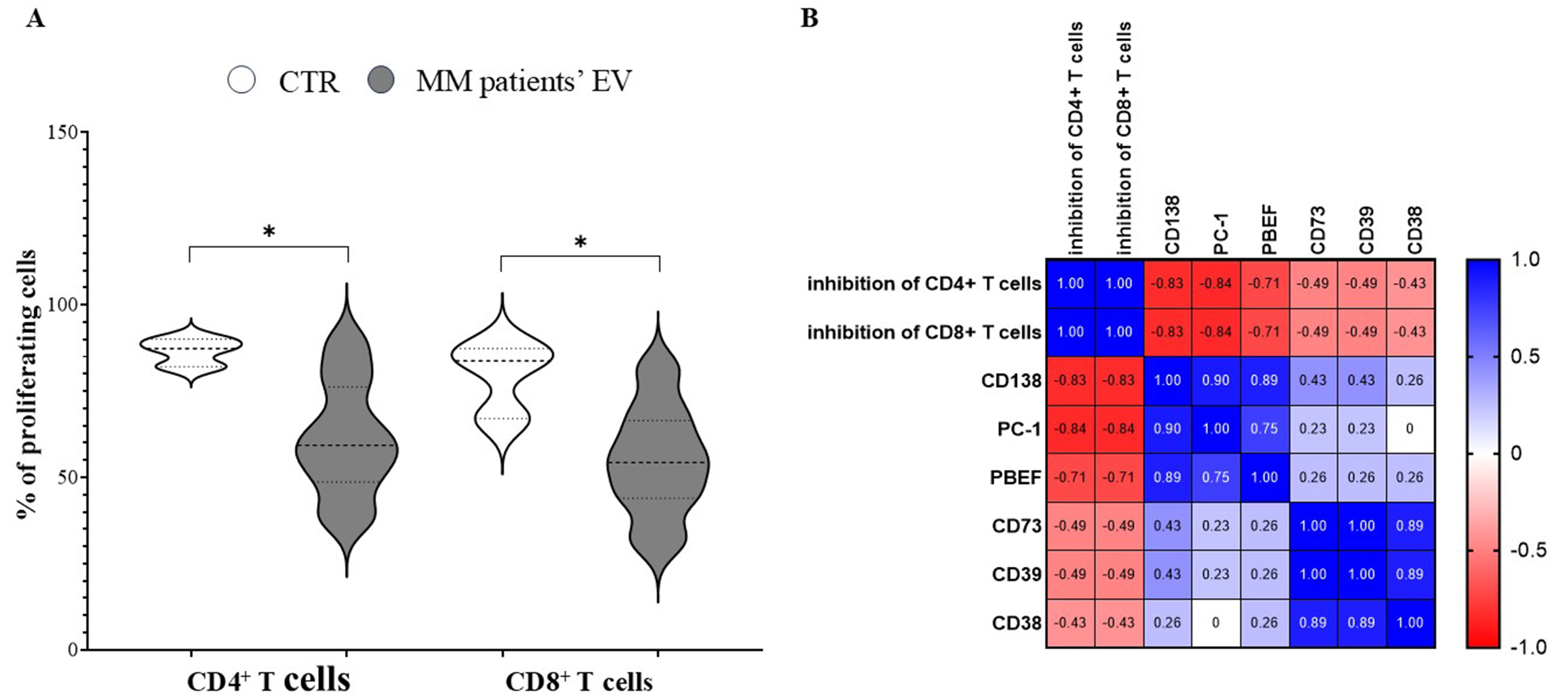
Figure 4. Immune-regulatory properties of MM-derived EVs. (A) Violin plots indicate the percentage of CD4+ and CD8+ T cells that proliferated after SEB stimulation, in the presence (grey profiles) or absence (white profiles) of MM-derived EVs. Profile spikes indicate maximum and minimum values, whereas dotted horizontal lines indicate quartiles and medians. Statistically significant differences were calculated using the Mann-Whitney test (*P ≤ 0.05) and are indicated by an asterisk. Correlations between different parameters, calculated by linear regression, are shown in the correlogram in (B) including (i) inhibition of CD4+ or CD8+ T cell proliferation and (ii) expression of different molecules on EVs. Blue indicates positive correlations (between 0 and 1), whereas red indicates inverse correlations (between 0 and -1). CTR: Control; MM: multiple myeloma; EVs: extracellular vesicles; SEB: staphylococcal enterotoxin B; PBEF: pre-B-cell colony-enhancing factor; PC-1: plasma cell antigen-1 (ectonucleotide pyrophosphatase/phosphodiesterase 1, ENPP1); CD: cluster of differentiation.
Finally, we assessed potential correlation between the down-modulation of T-cell proliferation and expression of surface markers on MM patients’ EVs (i.e., CD138, NAMPT/PBEF1, CD38, CD39, CD73, and CD203a). As shown in the correlogram in Figure 4B, inhibition of CD4⁺ and CD8+ T-lymphocyte proliferation correlated with the expression of CD138, CD203a, and NAMPT/PBEF1 (PBEF), and, to a lesser extent, with CD38, CD39, and CD73 on MM-derived EVs. However, statistically significant correlations were observed only between CD4⁺ and CD8+ T-cell proliferation and the expression of CD138 (P = 0.029) and CD203a (P = 0.0022) on EVs.
DISCUSSION
In recent years, several studies have acknowledged the involvement of EVs in tumor progression and tumorigenesis both in solid and hematological malignancies[26,27]. In addition, EVs are used as biomarkers for diagnostic purposes and for monitoring the course of different tumors. They have also been proposed as potential targets for novel therapeutic strategies[28,29]. Indeed, tumor-derived EVs can modulate the tumor microenvironment by mediating intercellular communication, thereby promoting immune escape, metastatic dissemination and resistance to therapy[24,27]. Importantly, EVs may provide insights into disease states and serve as diagnostic tools for early cancer detection, as they reflect gene mutations as well as phenotypic and functional features of their cells of origin, enabling minimally invasive tracking of disease progression[30-32].
For this reason, the characterization of circulating tumor-derived EVs in the PB as a form of liquid biopsy has attracted considerable interest for diagnostic and therapeutic purposes in recent years[30,33-35]. Accordingly, several studies have examined EVs in the PB of patients with solid and hematological malignancies, characterizing their role in tumor progression as well as their diagnostic and prognostic value[33,36-41]. More recently, Reale and coworkers described different methods for EVs isolation and characterization from the PB of patients with MM, highlighting their relevance for diagnostic and monitoring purposes[42]. However, detailed phenotypic and functional characterization of these EVs remains limited. The present study addresses this gap.
Here we report that PB samples from MM patients, compared with those from HD, contained an increased concentration of EVs, while displaying a similar size distribution. Increased levels of circulating EVs compared with healthy subjects have already been reported in patients with lung cancer, where they correlate with disease stage[43]. We also found that EVs obtained from MM patients’ PB samples displayed higher expression levels of CD138, NAMPT/PBEF1, CD38, and CD73 than those from HDs. The increased expression of CD138 on EVs from MM patients confirms that tumor-derived, CD138-expressing EVs are detectable in the PB of MM patients, and not only in the BM niche, as previously reported[21]. Nonetheless, the detection of CD138 on EVs from HDs, although at lower levels, was not entirely unexpected. Indeed, CD138 can be detected on normal epithelial cells, normal PCs, and other healthy tissues[44,45], and its expression can be increased during inflammation[46] or in autoimmune and inflammatory diseases[47,48].
The finding that NAMPT/PBEF1 is expressed at higher levels in EVs from MM patients than in those from HDs is novel and of particular interest. NAMPT/PBEF1 is a rate-limiting enzyme involved in NAD+ biosynthesis, and its expression has been reported in MM, where is crucial for tumor cell growth and osteoclast activity[18]. More importantly, NAMPT/PBEF1 expression has been shown to correlate with disease progression[49]. In addition, it has been proposed both as a therapeutic target[50] and as a biomarker of chemotherapy efficacy in patients with NDMM, particularly in combination with nicotinate phosphoribosyltransferase (NAPRT)[51]. Among the adenosinergic pathways, we observed significantly higher expression of CD38, which belongs to the non-canonical pathway, and of CD73, which represents a functional bridge between the canonical and non-canonical pathways. These data are consistent with our previous observations on EVs obtained from BM samples of MM patients[21]. Accordingly, EVs potentially capable of producing the immunosuppressive molecule ADO from different substrates, including ATP and NAD+, are detectable in the PB of MM patients and not only within the BM microenvironment. Furthermore, we demonstrated that CD138+ EVs, which are mostly derived from malignant PCs (although the contribution of CD138+ healthy cells cannot be excluded), expressed significantly higher levels of NAMPT/PBEF1, CD38, CD39, and CD73. These data suggested that those EVs display an increased capacity for NAD+ and ATP metabolism leading to ADO production, as compared with EVs derived from healthy cells. Again, this feature mirrors what has been observed in EVs isolated from BM samples of MM patients[21]. During MM progression, the BM niche becomes enriched in extracellular nucleotides (e.g., ATP and NAD⁺), as well as dinucleotides such as cyclic Guanosine monophosphate (GMP)-AMP (cGAMP). It has been reported that both mono- and di-nucleotides may contribute to the establishment of immune tolerance[52], through their metabolization by ectoenzymes expressed by BM resident cells, such as CD38, CD203a, and CD73. The resulting enzymatic reactions trigger intracellular calcium flux signaling and drive the production of extracellular ADO, thereby linking purinergic metabolism to immunosuppressive processes[4].
In this scenario, the interplay between the DNA damage response and purinergic metabolism remodeling is increasingly recognized as a critical determinant of MM adaptation to therapy. Genotoxic stress induces the release of ATP, NAD⁺, and cGAMP, which are rapidly processed by ectonucleotidases in the MM microenvironment. Among these ectonucelotidases, CD203a acts as a central regulator by converting ATP and NAD⁺ to AMP[53]. Thus, CD203a activity fuels the non-canonical adenosinergic pathway independently of the enzymatic function of CD39[9]. Furthermore, CD203a can suppress cGAS-Stimulator of Interferon Genes (STING)-mediated innate immune activation by degrading extracellular cGAMP[53]. In MM, PCs and BM stromal cells frequently upregulate CD203a, enabling efficient conversion of therapy-induced nucleotides into AMP and sustaining CD73 dependent ADO accumulation[54]. The combined enhancement of ADO signaling and the attenuation of STING responses promotes an immunosuppressive microenvironment that supports MM cell survival and contributes to therapeutic resistance.
Data from the functional studies reported here indicate that EVs obtained from MM patients’ PB samples may exert immune-suppressive activities, as evidenced by their ability to dampen T-cell proliferation in vitro in both helper and cytotoxic T lymphocytes. Previous studies on EVs derived from patients with MM have reported inhibitory effects on osteogenesis and angiogenesis in vitro[55,56].
Several studies support the concept that EVs in MM act as key mediators that reprogram the BM microenvironment into an immunosuppressive niche, thus facilitating tumor progression through multiple, parallel mechanisms. One of the primary mechanisms involves the polarization of macrophages toward a pro-tumorigenic M2 phenotype. Exosomal miR-let-7c derived from mesenchymal stem cells drives this M2 shift, while MM-derived exosomes specifically increase the expression of M2-associated markers such as Arginase (Arg)-1, interleukin (IL)-10, and CD206 in resting macrophages[57,58]. Furthermore, tumor-derived small EVs induce the expression of Programmed Death-Ligand (PD-L)-1 and pro-inflammatory cytokines such as IL-6 in macrophages by activating the Signal Transducer and Activator of Transcription (STAT) 3 and Toll-like receptor (TLR) 4/Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathways[59]. Beyond macrophages, EVs are instrumental in the expansion and activation of myeloid-derived suppressor cells (MDSCs). In this context, Bone Marrow Stromal Cells (BMSC)-derived exosomes are internalized by MDSCs, directly promoting their survival through activation of STAT1 and STAT3 signaling and upregulation of the anti-apoptotic proteins B-cell lymphoma-extra large (Bcl-xL) and Myeloid Cell Leukemia-1(Mcl-1)[60]. T-cell activity is likewise severely compromised through multiple EV-mediated pathways. Hagiwara et al. reported that EVs derived from MM promote T-cell exhaustion by upregulating immune checkpoint markers such as PD-1, T-cell immunoreceptor with Ig and ITIM domains (TIGIT), and Lymphocyte-activation gene (LAG) 3 on CD8+ T cells, a process largely driven by Sphingosine kinase (SPHK) 1/Sphingosine-1-phosphate (S1P) signaling[61]. Moreover, conditioning of the BM microenvironment with these vesicles increases PD-1 expression specifically on CD4+ T cells[62].
Here, we also report that circulating EVs in PB are equipped with complete adenosinergic pathways (CD38, CD39, CD203a, and CD73). This feature enables them to produce the immunosuppressive molecule ADO. Thus, circulating EVs are capable of dampening both CD4+ and CD8+ T-cell proliferation at sites far from the primary tumor. Natural killer (NK) cells are also targeted by EV-mediated mechanisms, since chronic exposure to EVs carrying the MHC class I polypeptide-related sequence A (MICA) ligand leads to downmodulation and degradation of the Natural Killer Group (NKG) 2D receptor and can even trigger “fratricide” when MICA is transferred directly from vesicles to the NK-cell surface[63]. In addition, exosomal lncRNA Nuclear Paraspeckle Assembly Transcript (NEAT) 1 inhibits NK-cell activity and cytokine production by downregulating Pre-B-cell leukemia transcription factor (PBX) 1 through an Enhancer of zeste homolog (EZH) 2-mediated axis[49]. Finally, MM EVs are enriched in Complement Component 4 Binding Protein Alpha (C4BPA), which helps tumor cells evade the host complement system[64], and phosphatidylserine-positive vesicles contribute to a highly immunosuppressive state[65].
We have further recently demonstrated that EVs obtained from BM samples of MM patients regulate multiple immune cell functions, including T-cell proliferation and cytokine release. Those effects are partly mediated by the immune checkpoints Human Leukocyte Antigen (HLA)-G and PD-L1 expressed on their surface[66]. Moreover, EVs from BM samples of MM patients express functional adenosinergic ectoenzymes, paralleled by the production of ADO from ATP and NAD+[21]. Based on these observations, we previously hypothesized that EVs released within the BM microenvironment may modulate local anti-tumor immune responses. Furthermore, those EVs may arrive in the PB and, from there, they reach other BM niches or other organs (see Graphical Abstract). Here we provide the first evidence that EVs detected in MM patients’ PB samples have potential immunomodulatory effects. It is conceivable that these EVs may recirculate through the blood stream to perform these modulatory activities on immune cells at distant sites, rather than in the PB. Downregulation of T-cell proliferation in vitro positively and significantly correlated with the expression of CD138 and CD203a/PC-1 on EVs, suggesting that these effects can be attributed, at least in part, to the presence of tumor-derived EVs expressing a functional non-canonical adenosinergic pathway. Nonetheless, these findings require validation using specific ectoenzyme inhibitors, and we cannot exclude the contribution of additional immune-suppressive mechanisms. Further studies will therefore be necessary to validate the data here presented. Nonetheless, we provide the first evidence that immune-regulatory EVs derived from MM cells are clearly present in the PB of MM patients. Those EVs may be involved in tumor-derived immune-suppression in the periphery and, presumably, in tumor progression.
This study has limitations, including the relatively small number of MM patients analyzed. Although the use of primary patient samples inevitably introduces a degree of clinical heterogeneity, the application of strict inclusion criteria, uniform pre-analytical processing, and standardized functional assays mitigates this limitation. Nevertheless, future studies based on larger and clinically stratified cohorts will be essential to dissect the relative contribution of disease stage, comorbidities, and prior treatments to EV-mediated immune modulation in MM. Another limitation is the lack of experiments employing specific inhibitors of adenosinergic ectoenzymes to directly assess their role in the immune-regulatory properties of MM-derived EVs. Despite these limitations, our findings, together with results from other groups, underscore the relevance of EVs found in the PB of MM patients. Circulating CD138+ microparticle levels accurately reflect tumor burden, often signaling early relapse weeks before symptoms or standard tests show any change[65]. The proteomic content of EVs, specifically the enrichment of CD44, is a key indicator of corticosteroid resistance and a predictor of shorter overall survival[67]. Beyond tracking the tumor itself, MM-EVs inhibit bone formation through transfer of molecules like miR-103a-3p, providing a precise biomarker for the diagnosis of bone lesions[68]. EVs may also enable real-time monitoring of treatment failure, as the presence of P-glycoprotein and phosphatidylserine on microparticles correlates with multidrug resistance and aggressive disease progression[65]. Finally, it has been established that a high EV cargo ratio (> 0.6 × 108 particles) is a significant prognostic parameter associated with immune dysfunction and lower survival rates, supporting a more personalized approach to long-term clinical management[64]. In conclusion, PB EVs may represent a valuable tool for the diagnosis and monitoring of MM, potentially overcoming some limitations of invasive BM procedures; however, additional studies are required to confirm these hypotheses.
DECLARATIONS
Acknowledgments
We deeply thank Laura McLean, Lecturer at the Department of Cultures, Politics and Society - University of Torino. Graphical Abstract was created withBioRender. Morandi, F. (2026) https://BioRender.com/crjsmyi.
Authors’ contributions
Analyzed data and wrote the manuscript: Airoldi I
Provided samples from patients, analyzed data, and wrote the manuscript: Soncini D
Performed experiments, analyzed data, and wrote the manuscript: Marimpietri D
Provided reagents (purified monoclonal antibodies) and wrote the manuscript: Malavasi F, Horenstein AL
Analyzed clinical data: Garibotto M, Cea M
Wrote the manuscript: Cea M
Performed experiments, analyzed data and wrote the manuscript: Morandi F
Availability of data and materials
Raw data are available upon reasonable request from the corresponding author.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported by the “Research to Care” grant from Sanofi to Morandi F, Italian Ministry of Health (Ricerca Corrente 2024 n°2042 and 5 × 1,000 project 5M-2022-23685490), Associazione Italiana per la Ricerca sul Cancro (AIRC, IG #23438, to Cea M), International Myeloma Society (IMS) and the Paula and Rodger Riney Foundation Translational Research Award 2023, to Cea M.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Human samples were obtained in accordance with the Declaration of Helsinki. Written informed consent to participate in the study was obtained from all subjects. The study was approved by the Ethics Committee (CER Liguria: 409/2021 - DB ID 11602).
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
3. Costa F, Dalla Palma B, Giuliani N. CD38 expression by myeloma cells and its role in the context of bone marrow microenvironment: modulation by therapeutic agents. Cells. 2019;8:1632.
4. Quarona V, Ferri V, Chillemi A, et al. Unraveling the contribution of ectoenzymes to myeloma life and survival in the bone marrow niche. Ann N Y Acad Sci. 2015;1335:10-22.
5. Kaczmarek E, Koziak K, Sévigny J, et al. Identification and characterization of CD39/vascular ATP diphosphohydrolase. J Biol Chem. 1996;271:33116-22.
6. Zimmermann H. 5-Nucleotidase: molecular structure and functional aspects. Biochem J. 1992;285:345-65.
7. Lennon PF, Taylor CT, Stahl GL, Colgan SP. Neutrophil-derived 5’-adenosine monophosphate promotes endothelial barrier function via CD73-mediated conversion to adenosine and endothelial A2B receptor activation. J Exp Med. 1998;188:1433-43.
8. Goding JW, Grobben B, Slegers H. Physiological and pathophysiological functions of the ecto-nucleotide pyrophosphatase/phosphodiesterase family. Biochim Biophys Acta. 2003;1638:1-19.
9. Horenstein AL, Chillemi A, Zaccarello G, et al. A CD38/CD203a/CD73 ectoenzymatic pathway independent of CD39 drives a novel adenosinergic loop in human T lymphocytes. Oncoimmunology. 2013;2:e26246.
10. Aliagas E, Vidal A, Texidó L, Ponce J, Condom E, Martín-Satué M. High expression of ecto-nucleotidases CD39 and CD73 in human endometrial tumors. Mediators Inflamm. 2014;2014:509027.
11. Deterre P, Gelman L, Gary-Gouy H, et al. Coordinated regulation in human T cells of nucleotide-hydrolyzing ecto-enzymatic activities, including CD38 and PC-1. Possible role in the recycling of nicotinamide adenine dinucleotide metabolites. J Immunol. 1996;157:1381-8.
12. Horenstein AL, Quarona V, Toscani D, et al. Adenosine generated in the bone marrow niche through a CD38-mediated pathway correlates with progression of human myeloma. Mol Med. 2016;22:694-704.
13. Sharif T, Ahn DG, Liu RZ, et al. The NAD+ salvage pathway modulates cancer cell viability via p73. Cell Death Differ. 2016;23:669-80.
14. Venter G, Oerlemans FT, Willemse M, Wijers M, Fransen JA, Wieringa B. NAMPT-mediated salvage synthesis of NAD+ controls morphofunctional changes of macrophages. PLoS One. 2014;9:e97378.
15. Yaku K, Okabe K, Hikosaka K, Nakagawa T. NAD metabolism in cancer therapeutics. Front Oncol. 2018;8:622.
16. Yong J, Cai S, Zeng Z. Targeting NAD+ metabolism: dual roles in cancer treatment. Front Immunol. 2023;14:1269896.
17. Reddy PS, Umesh S, Thota B, et al. PBEF1/NAmPRTase/Visfatin: a potential malignant astrocytoma/glioblastoma serum marker with prognostic value. Cancer Biol Ther. 2008;7:663-8.
18. Venkateshaiah SU, Khan S, Ling W, et al. NAMPT/PBEF1 enzymatic activity is indispensable for myeloma cell growth and osteoclast activity. Exp Hematol. 2013;41:547-57.e2.
19. Chu M, Rong J, Wang Y, et al. Strong association of the polymorphisms in PBEF1 and knee OA risk: a two-stage population-based study in China. Sci Rep. 2016;6:19094.
20. Colombo G, Clemente N, Zito A, et al. Neutralization of extracellular NAMPT (nicotinamide phosphoribosyltransferase) ameliorates experimental murine colitis. J Mol Med. 2020;98:595-612.
21. Morandi F, Marimpietri D, Horenstein AL, et al. Microvesicles released from multiple myeloma cells are equipped with ectoenzymes belonging to canonical and non-canonical adenosinergic pathways and produce adenosine from ATP and NAD+. Oncoimmunology. 2018;7:e1458809.
22. Castillo-Peña A, Molina-Pinelo S. Landscape of tumor and immune system cells-derived exosomes in lung cancer: mediators of antitumor immunity regulation. Front Immunol. 2023;14:1279495.
23. Giusti I, Poppa G, Di Fazio G, D’Ascenzo S, Dolo V. Metastatic dissemination: role of tumor-derived extracellular vesicles and their use as clinical biomarkers. Int J Mol Sci. 2023;24:9590.
24. Mitchell MI, Loudig O. Communicator extraordinaire: extracellular vesicles in the tumor microenvironment are essential local and long-distance mediators of cancer metastasis. Biomedicines. 2023;11:2534.
25. Wang S, Sun J, Dastgheyb RM, Li Z. Tumor-derived extracellular vesicles modulate innate immune responses to affect tumor progression. Front Immunol. 2022;13:1045624.
26. Xu R, Rai A, Chen M, Suwakulsiri W, Greening DW, Simpson RJ. Extracellular vesicles in cancer - implications for future improvements in cancer care. Nat Rev Clin Oncol. 2018;15:617-38.
27. Van Morckhoven D, Dubois N, Bron D, Meuleman N, Lagneaux L, Stamatopoulos B. Extracellular vesicles in hematological malignancies: EV-dence for reshaping the tumoral microenvironment. Front Immunol. 2023;14:1265969.
28. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024;9:27.
29. Urabe F, Kosaka N, Ito K, Kimura T, Egawa S, Ochiya T. Extracellular vesicles as biomarkers and therapeutic targets for cancer. Am J Physiol Cell Physiol. 2020;318:C29-39.
30. Hao X, Liu Z, Ma F, et al. Exosome-based liquid biopsy in early screening and diagnosis of cancers. Dose Response. 2025;23:15593258251344480.
31. Santamaria S, Cardinali B, Rovere M, et al. New insight in early detection and precision medicine in small cell lung cancer: liquid biopsy as innovative clinical tool. Crit Rev Clin Lab Sci. 2025;62:404-28.
32. Thomas Junior DS, Chai J, Lu YJ. The development and applications of circulating tumour cells, circulating tumour DNA and other emerging biomarkers for early cancer detection. Explor Target Antitumor Ther. 2025;6:1002314.
34. Izhar M, Ahmad Z, Moazzam M, Jader A. Targeted liquid biopsy for brain tumors. J Liq Biopsy. 2024;6:100170.
35. De Rosa C, Amato L, Ariano A, et al. Novel applications of liquid Biopsy: comprehensive methodology for circulating biomarker exploration in peripheral blood. J Liq Biopsy. 2025;9:100307.
36. Benecke L, Chiang DM, Ebnoether E, Pfaffl MW, Muller L. Isolation and analysis of tumor‑derived extracellular vesicles from head and neck squamous cell carcinoma plasma by galectin‑based glycan recognition particles. Int J Oncol. 2022;61:133.
37. Zebrowska A, Widlak P, Whiteside T, Pietrowska M. Signaling of tumor-derived sEV impacts melanoma progression. Int J Mol Sci. 2020;21:5066.
38. Rappa G, Puglisi C, Santos MF, Forte S, Memeo L, Lorico A. Extracellular vesicles from thyroid carcinoma: the new frontier of liquid biopsy. Int J Mol Sci. 2019;20:1114.
39. Sun Y, Xing L, Luo J, et al. A pro-metastatic derivatives eliminator for in vivo dual-removal of circulating tumor cells and tumor-derived exosomes impedes their biodistribution into distant organs. Adv Sci. 2023;10:e2304287.
40. Kontopoulou E, Strachan S, Reinhardt K, et al. Evaluation of dsDNA from extracellular vesicles (EVs) in pediatric AML diagnostics. Ann Hematol. 2020;99:459-75.
41. Döring K, Malinova V, Bettag C, et al. The diagnostic potential of extracellular vesicles derived from the blood plasma of glioblastoma patients. In Vivo. 2024;38:2735-9.
42. Reale A, Khong T, Xu R, et al. Human plasma extracellular vesicle isolation and proteomic characterization for the optimization of liquid biopsy in multiple myeloma. Methods Mol Biol. 2021;2261:151-91.
43. Choi BH, Quan YH, Rho J, et al. Levels of extracellular vesicles in pulmonary and peripheral blood correlate with stages of lung cancer patients. World J Surg. 2020;44:3522-9.
44. Kind S, Merenkow C, Büscheck F, et al. Prevalence of syndecan-1 (CD138) expression in different kinds of human tumors and normal tissues. Dis Markers. 2019;2019:4928315.
45. Palaiologou M, Delladetsima I, Tiniakos D. CD138 (syndecan-1) expression in health and disease. Histol Histopathol. 2014;29:177-89.
46. Diab L, Al Kattar S, Oueini N, et al. Syndecan-1: a key player in health and disease. Immunogenetics. 2024;77:9.
47. Koliakou E, Eleni MM, Koumentakou I, et al. Altered distribution and expression of syndecan-1 and -4 as an additional hallmark in psoriasis. Int J Mol Sci. 2022;23:6511.
48. Luo L, Feng S, Wu Y, Su Y, Jing F, Yi Q. Serum levels of syndecan-1 in patients with Kawasaki disease. Pediatr Infect Dis J. 2019;38:89-94.
49. Expression of NAMPT in Patients with Multiple Myeloma and Its Correlation with Clinical Manifestation. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2023;31:769-76. (in Chinese) Available from: https://jglobal.jst.go.jp/en/detail?JGLOBAL_ID=202302273983252412. [Last accessed on 15 Apr 2026].
50. Bong IP, Ng CC, Fakiruddin SK, Lim MN, Zakaria Z. Small interfering RNA-mediated silencing of nicotinamide phosphoribosyltransferase (NAMPT) and lysosomal trafficking regulator (LYST) induce growth inhibition and apoptosis in human multiple myeloma cells: a preliminary study. Bosn J Basic Med Sci. 2016;16:268-75.
51. Soncini D, Becherini P, Ladisa F, et al. NAD+ metabolism restriction boosts high-dose melphalan efficacy in patients with multiple myeloma. Blood Adv. 2025;9:1024-39.
52. Horenstein AL, Morandi F, Bracci C, Pistoia V, Malavasi F. Functional insights into nucleotide-metabolizing ectoenzymes expressed by bone marrow-resident cells in patients with multiple myeloma. Immunol Lett. 2019;205:40-50.
53. Stagg J, Golden E, Wennerberg E, Demaria S. The interplay between the DNA damage response and ectonucleotidases modulates tumor response to therapy. Sci Immunol. 2023;8:eabq3015.
54. Morandi F, Horenstein AL, Costa F, Giuliani N, Pistoia V, Malavasi F. CD38: a target for immunotherapeutic approaches in multiple myeloma. Front Immunol. 2018;9:2722.
55. Guo HM, Sun L, Yang L, Liu XJ, Nie ZY, Luo JM. Microvesicles shed from bortezomib-treated or lenalidomide-treated human myeloma cells inhibit angiogenesis in vitro. Oncol Rep. 2018;39:2873-80.
56. Zhang L, Lei Q, Wang H, et al. Tumor-derived extracellular vesicles inhibit osteogenesis and exacerbate myeloma bone disease. Theranostics. 2019;9:196-209.
57. Razi B, Soleimani M, Soufi-Zomorrod M, Adeli A, Davoudi N. Multiple myeloma cell-derived exosomes promote favorable tumor functional performance by polarizing macrophages toward M2-like cells. APMIS. 2023;131:381-93.
58. Tian X, Sun M, Wu H, et al. Exosome-derived miR-let-7c promotes angiogenesis in multiple myeloma by polarizing M2 macrophages in the bone marrow microenvironment. Leuk Res. 2021;105:106566.
59. Pucci M, Raimondo S, Urzì O, et al. Tumor-derived small extracellular vesicles induce pro-inflammatory cytokine expression and PD-L1 regulation in M0 macrophages via IL-6/STAT3 and TLR4 signaling pathways. Int J Mol Sci. 2021;22:12118.
60. Wang J, De Veirman K, De Beule N, et al. The bone marrow microenvironment enhances multiple myeloma progression by exosome-mediated activation of myeloid-derived suppressor cells. Oncotarget. 2015;6:43992-4004.
61. Hagiwara S, Ri M, Ebina T, et al. Immunosuppressive effects of multiple myeloma-derived extracellular vesicles through T cell exhaustion. Cancer Sci. 2025;116:1861-70.
62. Lopes R, Caetano J, Barahona F, et al. Multiple myeloma-derived extracellular vesicles modulate the bone marrow immune microenvironment. Front Immunol. 2022;13:909880.
63. Vulpis E, Loconte L, Peri A, et al. Impact on NK cell functions of acute versus chronic exposure to extracellular vesicle-associated MICA: dual role in cancer immunosurveillance. J Extracell Vesicles. 2022;11:e12176.
64. Ferreira BV, Carneiro EA, Pestana C, et al. Patient-derived extracellular vesicles proteins as new biomarkers in multiple myeloma - a real-world study. Front Oncol. 2022;12:860849.
65. Krishnan SR, Luk F, Brown RD, Suen H, Kwan Y, Bebawy M. Isolation of human CD138+ microparticles from the plasma of patients with multiple myeloma. Neoplasia. 2016;18:25-32.
66. Soncini D, Marimpietri D, Ladisa F, et al. Bone marrow-derived extracellular vesicles from multiple myeloma patients promote adaptive immune dysfunction via HLA-G, PD-1, and PD-L1. Front Immunol. 2025;16:1640168.
67. Harshman SW, Canella A, Ciarlariello PD, et al. Proteomic characterization of circulating extracellular vesicles identifies novel serum myeloma associated markers. J Proteomics. 2016;136:89-98.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].