
Glymphatic enhancement as a therapeutic strategy in Alzheimer’s disease: a comprehensive review of preclinical evidence and translational opportunities
 0
0 
Abstract
The glymphatic system is a brain-wide waste clearance network responsible for eliminating metabolic byproducts, including amyloid-beta (Aβ) and tau, two major pathological hallmarks of Alzheimer’s disease (AD). This review synthesizes current evidence indicating that glymphatic dysfunction, driven by structural and molecular impairments in meningeal lymphatic vessels, perivascular spaces, and aquaporin-4 (AQP4) channels, plays a critical role in the pathogenesis and progression of AD. Multiple contributing factors, including aging, sleep disturbances, and genetic susceptibility such as apolipoprotein E4 (APOE4), further aggravate glymphatic impairment, thereby promoting Aβ accumulation, neuroinflammation, and cognitive decline. Emerging therapeutic strategies aimed at restoring glymphatic function include pharmacological interventions, such as melatonin, cilostazol, and vascular endothelial growth factor C (VEGF-C), as well as traditional Chinese medicine approaches, including Borneol and Yuanzhi Powder, and neuromodulation-based therapies such as focused ultrasound and electroacupuncture. In addition, lifestyle interventions, particularly physical exercise and sleep optimization, may enhance cerebrospinal fluid (CSF) dynamics and preserve AQP4 polarization, thereby facilitating metabolic clearance. Preclinical studies have demonstrated that these interventions can reduce Aβ and tau deposition, suppress neuroinflammation, and improve cognition. However, substantial clinical validation remains necessary before these findings can be translated into effective therapeutic applications. Collectively, this review highlights the glymphatic system as a promising therapeutic target for AD and provides an integrated perspective linking mechanistic insights with emerging therapeutic strategies.
Keywords
INTRODUCTION
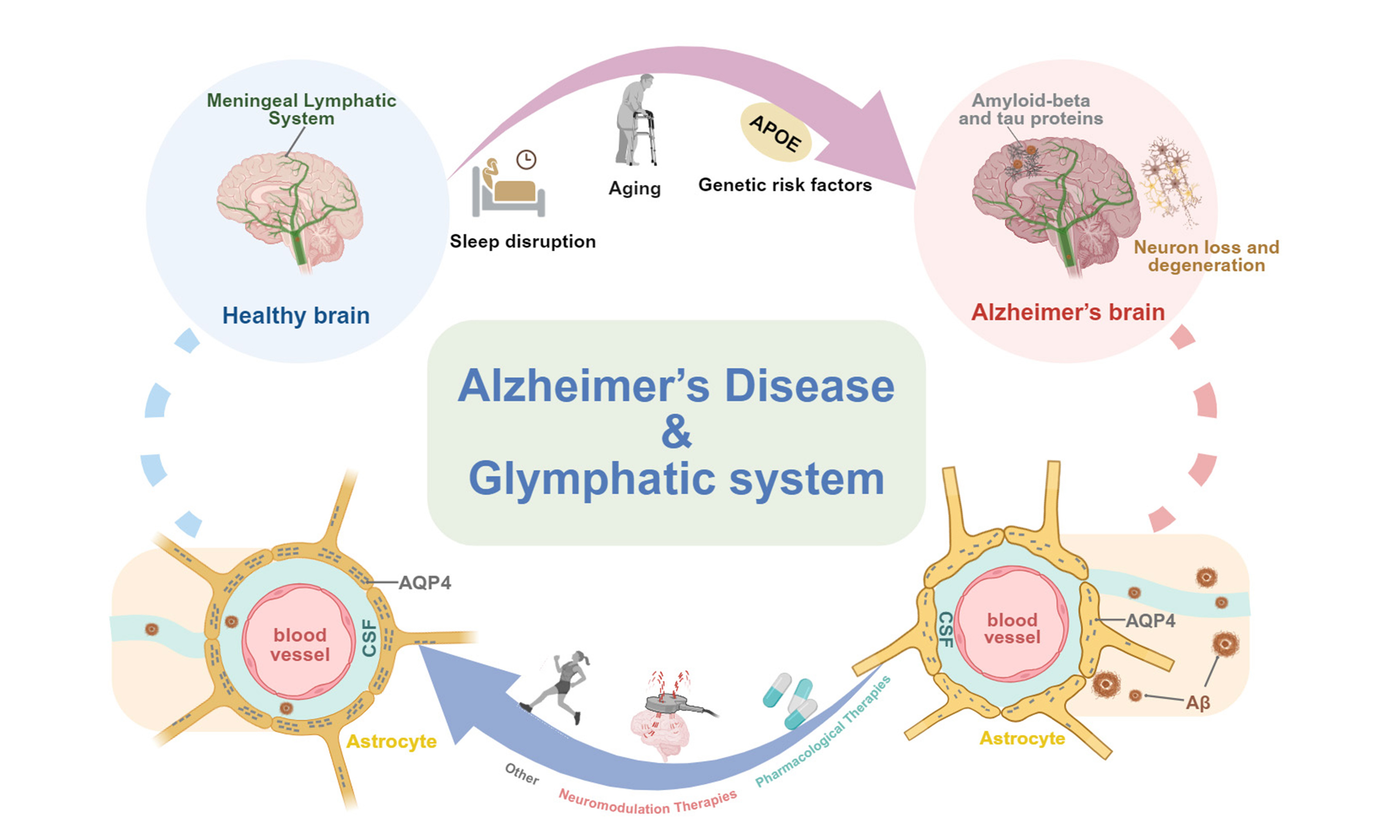
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder characterized by the accumulation of amyloid-beta (Aβ) plaques and tau neurofibrillary tangles, ultimately leading to cognitive impairment and dementia[1]. The discovery of the glymphatic system in 2012 identified a brain-wide waste clearance pathway that facilitates the removal of neurotoxic metabolites through perivascular channels, cerebrospinal fluid (CSF) circulation, and astrocytic aquaporin-4 (AQP4) water channels[2,3]. Increasing evidence indicates that glymphatic dysfunction, driven by structural and molecular impairments in meningeal lymphatic vessels (mLVs), perivascular spaces, and AQP4 mislocalization, contributes to the accumulation of Aβ and tau, exacerbates neuroinflammation and synaptic injury, and thereby accelerates AD progression[4].
Recent studies further suggest that restoration or enhancement of glymphatic function may represent a promising therapeutic strategy for AD. Preclinical evidence demonstrates that pharmacological agents, traditional Chinese medicine (TCM) formulations, neuromodulation approaches, and lifestyle interventions can improve glymphatic transport, reduce pathological protein aggregation, and mitigate cognitive deficits[5-11]. This review summarizes current advances in understanding glymphatic dysfunction as a central contributor to AD pathogenesis and critically evaluates emerging therapeutic strategies targeting this glymphatic system, highlighting opportunities and challenges for clinical translation.
METHODOLOGY
A systematic literature search was conducted in Medline, the Cochrane Library, Web of Science, SCOPUS, and PsycINFO from database inception through March 2026. Search terms related to AD (“Alzheimer’s disease” OR “Alzheimer disease”) were combined with terms associated with brain lymphatic clearance pathways (“glymphatic system” OR “meningeal lymphatic” OR “lymphatic drainage”) using Boolean operators and database-specific search syntax.
Two reviewers independently screened titles, abstracts, and full-text articles for eligibility. Studies were included if they: (1) were original preclinical or clinical investigations examining glymphatic or meningeal lymphatic function in AD; (2) evaluated interventions targeting these systems; and (3) were published as peer-reviewed articles in English. Editorials, conference abstracts, case reports, non-English publications, and studies lacking direct relevance to glymphatic or meningeal lymphatic mechanisms in AD were excluded. Data extraction was independently performed by two reviewers using a standardized form, which included study design, experimental model, intervention strategy, principal findings, and outcome measures. Any discrepancies between reviewers were resolved through consensus discussion or, when necessary, consultation with a fourth reviewer. Therapeutic strategies were subsequently classified according to their underlying mechanisms of action and potential clinical applicability. To ensure the inclusion of the most up-to-date evidence, the literature search was updated in March 2026.
THE GLYMPHATIC AND MENINGEAL LYMPHATIC SYSTEMS
The central nervous system (CNS) was historically considered devoid of a conventional lymphatic drainage system until the discovery of the glymphatic system in 2012 identified a specialized pathway responsible for cerebral solute clearance[2]. Subsequent studies further demonstrated the presence of mLVs, establishing that these structures function in concert with the glymphatic system to constitute an integrated brain lymphatic drainage network[3]. Major milestones in glymphatic system research relevant to AD are illustrated in Figure 1.
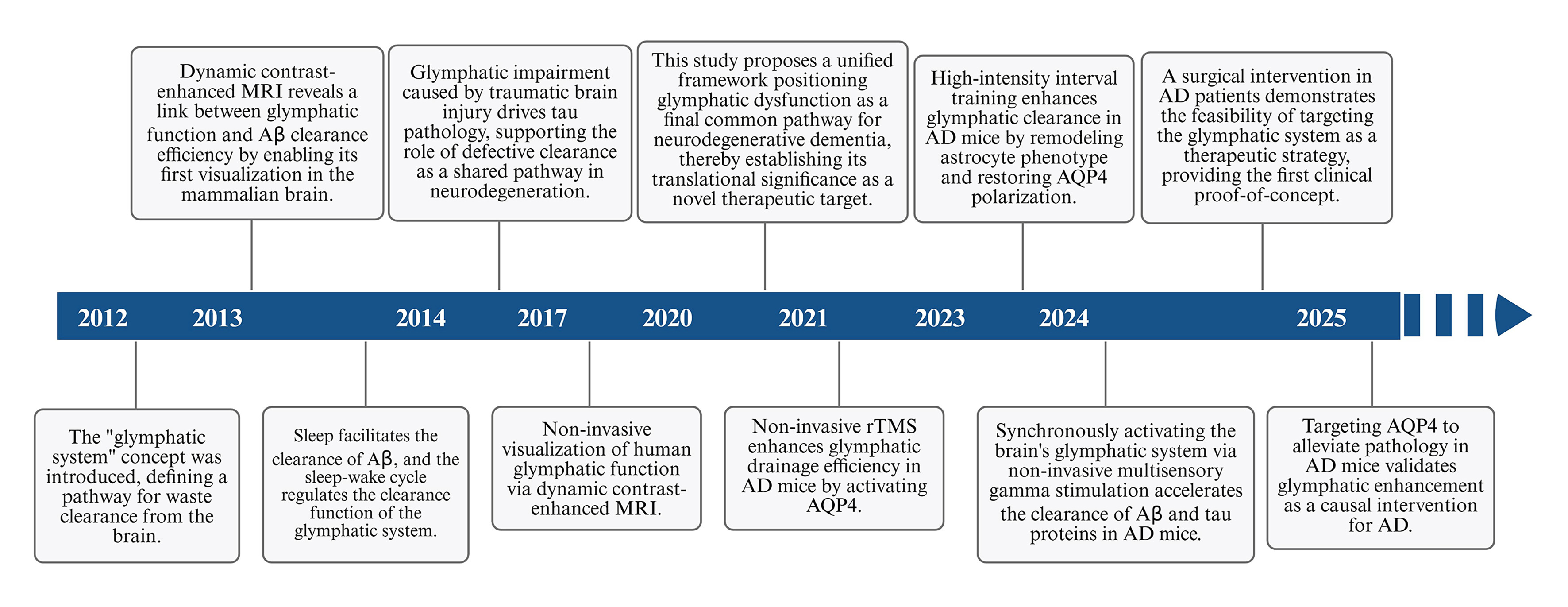
Figure 1. A timeline outlines key milestones in the research of the GS in AD. The discovery of the brain waste clearance system represented a major breakthrough in understanding the pathophysiology of AD. Since its identification, accumulating evidence has increasingly implicated glymphatic dysfunction as a critical contributor to AD development and progression. The timeline presented in Figure 1 illustrates key advances in this field and highlights the emerging importance of the glymphatic system in both the pathogenesis and theraputic targeting of AD. This figure was designed by the authors using BioRender. https://BioRender.com/ex09cg6. AD: Alzheimer’s Disease; GS: glymphatic system; MRI: magnetic resonance imaging; Aβ: amyloid-beta; AQP4: aquaporin-4; rTMS: repetitive transcranial magnetic stimulation.
The glymphatic system consists of perivascular spaces, astrocyte endfeet enriched with AQP4 water channels, CSF, and interstitial fluid (ISF)[12]. CSF enters the periarterial spaces primarily through arterial pulsation, a process facilitated by AQP4-mediated water transport, and subsequently circulates through the brain parenchyma under the influence of vascular and respiratory dynamics, thereby promoting the clearance of metabolic wastes products, including Aβ and tau proteins[13,14]. Waste-laden fluid is then drained through the perivenous spaces into the mLVs and ultimately transported to the deep cervical lymph nodes. Additional clearance pathways have also been identified along the cranial, olfactory, and spinal nerves [Figure 2][5].
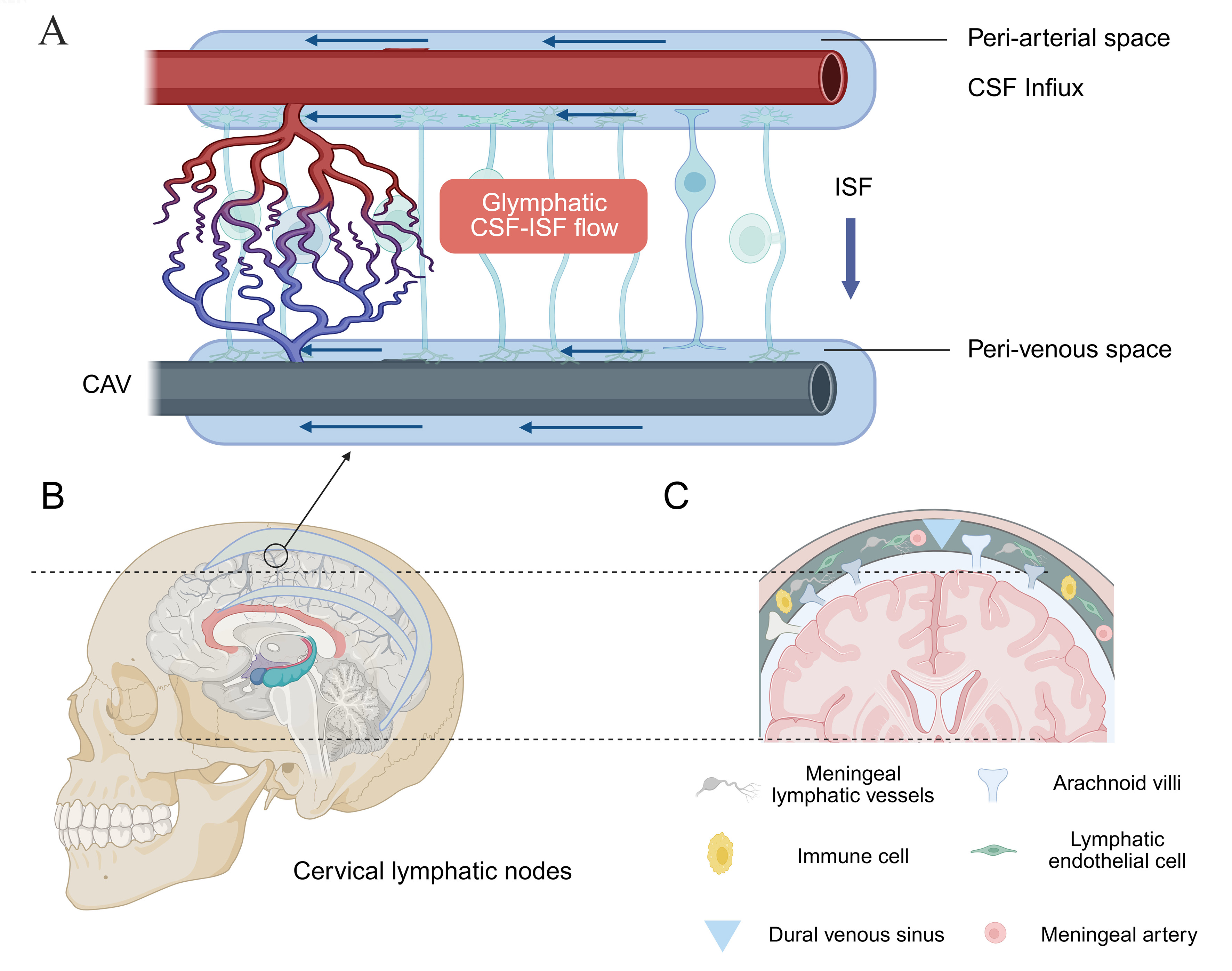
Figure 2. Glymphatic and meningeal lymphatic pathways. (A) CSF influx via periarterial spaces; (B) Metabolic waste drainage into the meningeal lymphatic vessels; (C) Outflow toward the deep cervical lymph nodes. This figure was designed by the authors using BioRender. https://BioRender.com/8449jme. CSF: Cerebrospinal fluid; ISF: interstitial fluid; CAV: cerebral arteriovenous.
Glymphatic activity is dynamically regulated by a wide range of physiological, behavioral, and pathological factors. Lifestyle-related variables, including physical exercise, sleep posture, dietary habits, and alcohol consumption, have all been shown to influence glymphatic transport efficiency[12,15-17]. Sleep markedly enhances glymphatic clearance, increasing solute removal by approximately two to threefold through expansion of the interstitial spaces, whereas aging impairs glymphatic function as a consequence of brain atrophy, reduced vascular compliance, and arterial stiffening[18]. In addition, cardiovascular status and lifestyle behaviors, particularly exercise, dietary patterns, and alcohol intake, further modulate glymphatic dynamics[15,16]. Current evidence suggests that the glymphatic system operates through four highly coordinated mechanisms. (1) arterial pulsation-driven CSF flow along periarterial spaces, maintained by parenchymal border macrophages that maintain extracellular matrix homeostasis through degradative remodeling; (2) sleep-associated neuronal activity generates rhythmic oscillations that facilitate ISF-CSF exchange; (3) neurovascular coupling regulates cerebral blood flow and thereby influences CSF movement within the brain parenchyma; (4) mLVs collect metabolic waste products and mediate their drainage to the deep cervical lymph nodes[3]. Furthermore, lymphatic valve-mediated unidirectional transport and vascular endothelial growth factor receptor 3 (VEGFR3) signaling are essential for maintaining glymphatic and lymphatic homeostasis[19].
mLVs in the dura mater drain a substantial proportion of CSF and facilitate clearance of neurotoxic waste via the deep cervical lymph nodes. Functionally, mLVs comprise specialized subtypes, including dorsal vessels involved in immune modulation, basal vessels that serve as the primary CSF drainage route, and ethmoidal mLVs. Critically, mLVs serve as key neuroimmune conduits by transporting CNS-derived antigens and immune cells, thereby mediating bidirectional central-peripheral immune communication. Dysfunction of the mLV system has been closely associated with aging and the pathogenesis of neurodegenerative disorders[20,21].
THE GLYMPHATIC AND MENINGEAL LYMPHATIC DYSFUNCTION IN AD
Emerging research implicates glymphatic system dysfunction as a key contributor to AD pathogenesis. In AD mice models, structural and functional impairments of mLVs, such as reduced vessel density, luminal narrowing, and diminished drainage capacity, compromise the clearance of neurotoxic proteins such as
Physiological factors affecting glymphatic and meningeal lymphatic function in AD
Aging is a primary driver of glymphatic function and represents the strongest risk factor for AD, with the prevalence of AD and other dementias approximately doubling every 5.5 years after the age of 60[24]. During aging, choroid plexus function declines, resulting in reduced CSF production, decreased arterial pulsatility, and slower CSF circulation, collectively impairing the efficient clearance of metabolic waste[25]. Consistently, aged mice exhibit significantly reduced CSF-lymphatic outflow compared with young counterparts[18]. In parallel, aging is associated with increased brain tissue stiffness, disrupted ISF dynamics, and structural alterations in mLVs, including reduced junctional integrity and impaired drainage capacity, all of which compromise glymphatic clearance[20,26].
Transcriptomic analyses further corroborate these findings, revealing age-dependent alterations in lymphatic endothelial cell gene expression profiles in mLVs[20]. The function of astrocytic water channels, such as AQP4, declines with age, further obstructing ISF flow. Additionally, chronic neuroinflammation progressively intensifies during aging, and pro-inflammatory mediators further disrupt glymphatic system integrity. Moreover, the age-related decline in microglial phagocytic capacity reduces the clearance of metabolic waste, thereby further compromising glymphatic system[27].
Sleep is a critical physiological regulator of glymphatic function. During natural sleep, the brain’s interstitial space expands, thereby facilitating CSF-ISF exchange and enhancing the clearance of Aβ. In sleep states, the exchange between CSF and ISF is markedly increased compared with wakefulness[18]. In mice, Aβ clearance approximately doubles during sleep compared with the awake state[18]. Conversely, sleep deprivation significantly reduces the clearance of various CSF metabolites in mice[18,28,29]. In humans, poor sleep quality is a well-established risk factor for cognitive decline and dementia[30]. In older adults, sleep is characterized by reduced duration and quality, increased fragmentation, impaired sleep maintenance, and alterations in rapid eye movement sleep architecture[31]. Both clinical and experimental evidence suggests that sleep regulates the extracellular clearance of AD-related proteins[32]. Preclinical studies demonstrate that interstitial Aβ levels decrease by approximately 25% during sleep, while tau clearance increases by up to 90%, underscoring the regulatory role of sleep in AD-related protein homeostasis[33].
Cardiac pulsation, respiration, and vascular pulsatility are key physiological drivers of glymphatic flow and subsequent meningeal lymphatic drainage. Studies have demonstrated that CSF convection within the glymphatic system is driven not only by cardiac pulsations but also by respiratory activity and very low-frequency vascular oscillations. These physiological forces shape CSF flow through unique spatiotemporal patterns and have enabled novel imaging approaches for the early detection and intervention of neurodegenerative diseases[34]. Clinical studies have shown that intrathecally administered MRI contrast agents can distribute throughout the entire brain, confirming direct communication between subarachnoid CSF and the perivascular spaces of brain tissue. These findings underscore the pivotal role of cardiac and respiratory influences in regulating glymphatic flow in humans[35].
Enhancement of glymphatic function in AD improves the clearance of Aβ and tau, attenuates neuroinflammation and oxidative stress, and restores ISF homeostasis. These effects contribute to the preservation of blood-brain barrier (BBB) integrity, maintenance of cerebrovascular function, and facilitation of neuronal recovery, thereby ultimately delaying cognitive decline. Accordingly, strategies aimed at optimizing glymphatic function, including sleep modulation, improvement of cardiovascular health, and pharmacological interventions, represent promising approaches to slow AD progression and improve clinical outcomes.
Immunological mechanisms underlying glymphatic and meningeal lymphatic dysfunction in AD
AD is characterized by profound immune dysfunction involving both central and peripheral immune systems. This pathological process is closely associated with impaired clearance function of the glymphatic system and mLVs. The primary manifestations include dysregulated activation of microglia and astrocytes[36], which contribute to chronic neuroinflammation, impaired clearance of pathological proteins, and subsequent neuronal injury[27]. In tauopathy mouse models and AD brains, tau pathology correlates with increased infiltration of cytotoxic T cells, which exacerbate neuroinflammation through the release of pro-inflammatory cytokines, such as interleukin-1β (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor-α (TNF-α). These cytokines suppress anti-inflammatory mediators such as IL-10, thereby exacerbating neuronal loss[37]. Activated cluster of differentiation 8-positive (CD8+) T cells further recruit interferon (IFN)-responsive microglia via chemokine signaling, establishing a feedforward inflammatory loop[38]. Infiltrating CD8+ T cells secrete cytotoxic effector molecules, including perforin and granzymes, which further amplify microglial activation and promote increased production of pro-inflammatory cytokines such as IFN-γ, IL-1β, and TNF-α. Critically, this inflammatory milieu disrupts Aβ clearance while promoting its aggregation, thereby directly linking immune dysregulation to amyloid pathology[39-41]. Additionally, pro-inflammatory cytokines alter synaptic and neuronal gene expression in APP/PS1 transgenic mice, thereby accelerating cognitive decline[42].
Beyond CD8+ T cells, CD4+ T cells also contribute to the progression of AD and regulate glymphatic and meningeal lymphatic drainage through various mechanisms, with their dysfunction increasing the risk of AD[27,43]. Immune-related factors, including pro-inflammatory cytokines, apolipoprotein E4 (APOE4), triggering receptor expressed on myeloid cells 2 (TREM2), and cluster of differentiation 22 (CD22), play critical roles in regulating neuroinflammation and lymphatic drainage. Dysregulation of these factors compromises BBB integrity, enabling peripheral immune cell infiltration and exacerbating neurodegeneration[27,44]. Impaired glymphatic and meningeal lymphatic drainage exacerbates Aβ accumulation and neuroinflammation, while Aβ deposits further obstruct glymphatic pathways, forming a self-reinforcing pathogenic loop that drives disease progression[45]. Chronic meningeal lymphatic dysfunction disrupts cortical excitatory-inhibitory balance via microglial IL-6 signaling, contributing to cognitive deficits, whereas restoration of lymphatic function reverses aging-related synaptic and behavioral impairments[46]. Besides, enhancement of lymphatic function [e.g., via vascular endothelial growth factor C (VEGF-C) therapy] enhances Aβ clearance and synergizes with anti-amyloid immunotherapies, highlighting combinatorial strategies targeting neurovascular-immune crosstalk[21,47]. Collectively, these findings elucidate lymphatic-immune interactions as critical therapeutic nodes for mitigating AD pathology [Figure 2].
The impact of AQP4 on glymphatic and lymphatic drainage in AD
AQP4, a water channel highly enriched at astrocyte endfeet, plays a pivotal role in maintaining fluid homeostasis and glymphatic function, thereby facilitating cerebral waste clearance. In AD, AQP4 loses its polarized localization at astrocyte endfeet and redistributes to non-perivascular regions. Although this redistribution may initially exert neuroprotective effects by promoting astrocyte plasticity and promoting the formation of protective glial barriers around Aβ plaques, persistent AQP4 mislocalization ultimately disrupts fluid dynamics and accelerates neuropathological changes[48]. Consistently, AQP4 deficiency markedly aggravates AD pathology. AQP4-null mice exhibit a 70% reduction in lymphatic drainage volume and a 55% decline in interstitial solute clearance, leading to enhanced Aβ accumulation and progressive cognitive impairment[2,49]. Collectively, these findings underscore the indispensable role of AQP4 in preventing Aβ-driven neurodegeneration.
Notably, AQP4 function declines with aging, further compromising glymphatic efficiency. In aged mice, Aβ clearance is reduced by approximately 40% compared to young mice, paralleled by impaired perivascular AQP4 polarization along cerebral arteries and diminished CSF-ISF exchange[25]. Similarly, in AD mouse models, Aβ deposition correlates with AQP4 redistribution away from astrocyte endfeet, disrupting water transport and ionic homeostasis[50]. Human studies further support these observations, demonstrating an age-dependent reductions in perivascular AQP4 localization. Such alterations may increase the susceptibility of the aging brain to pathological protein aggregation, particularly Aβ accumulation. Taken together, these observations identify AQP4 mislocalization as a critical contributor to glymphatic dysfunction and AD progression[51].
The impact of APOE on glymphatic and meningeal lymphatic drainage in AD
The APOE ε4 allele, the strongest genetic risk factor for late-onset AD, disrupts glymphatic and meningeal lymphatic drainage through multiple pathological mechanisms. APOE4 impairs Aβ clearance, compromises BBB integrity, induces cerebrovascular dysfunction, and exacerbates neuroinflammation and oxidative stress, collectively hindering CSF and ISF circulation[52-54]. In transgenic mouse models, APOE4 expression triggers BBB breakdown by activating inflammatory signaling pathways in pericytes, which are critical for maintaining vascular stability[55].
Accumulating evidence further suggests that APOE directly regulates glymphatic and lymphatic system function. One study found that the APOE4 genotype significantly enhances the association between glymphatic dysfunction, as assessed by the along the perivascular space (ALPS) index, and cerebral Aβ deposition. Specifically, a lower ALPS index correlated with a greater Aβ burden exclusively in APOE4 carriers, whereas no significant correlation was observed with tau pathology[56]. Additionally, ApoE derived from the choroid plexus and CSF enters the brain through the glymphatic pathway along perivascular spaces and is subsequently distributed to neurons. Notably, this distribution occurs in an isoform-dependent manner (apoE2 > apoE3 > apoE4) and is regulated by AQP4. Notably, sleep deprivation markedly disrupts this glymphatic transport process[57]. Furthermore, APOE4 has been hypothesized to induce premature shrinkage and sclerosis of mLVs, a pathological process termed meningeal lymphosclerosis, leading to lymphatic dysfunction. Impaired meningeal lymphatic drainage may reduce the clearance of Aβ, cellular debris, inflammatory mediators, and immune cells from the CNS, ultimately accelerating AD progression and cognitive decline. Supporting this hypothesis, RNA-Seq analyses have shown that induced pluripotent stem cells (iPSCs) carrying the APOE4 allele exhibit significantly reduced expression of genes associated with lymphatic endothelial cell markers and hereditary lymphedema-related pathways[52][Figure 3].
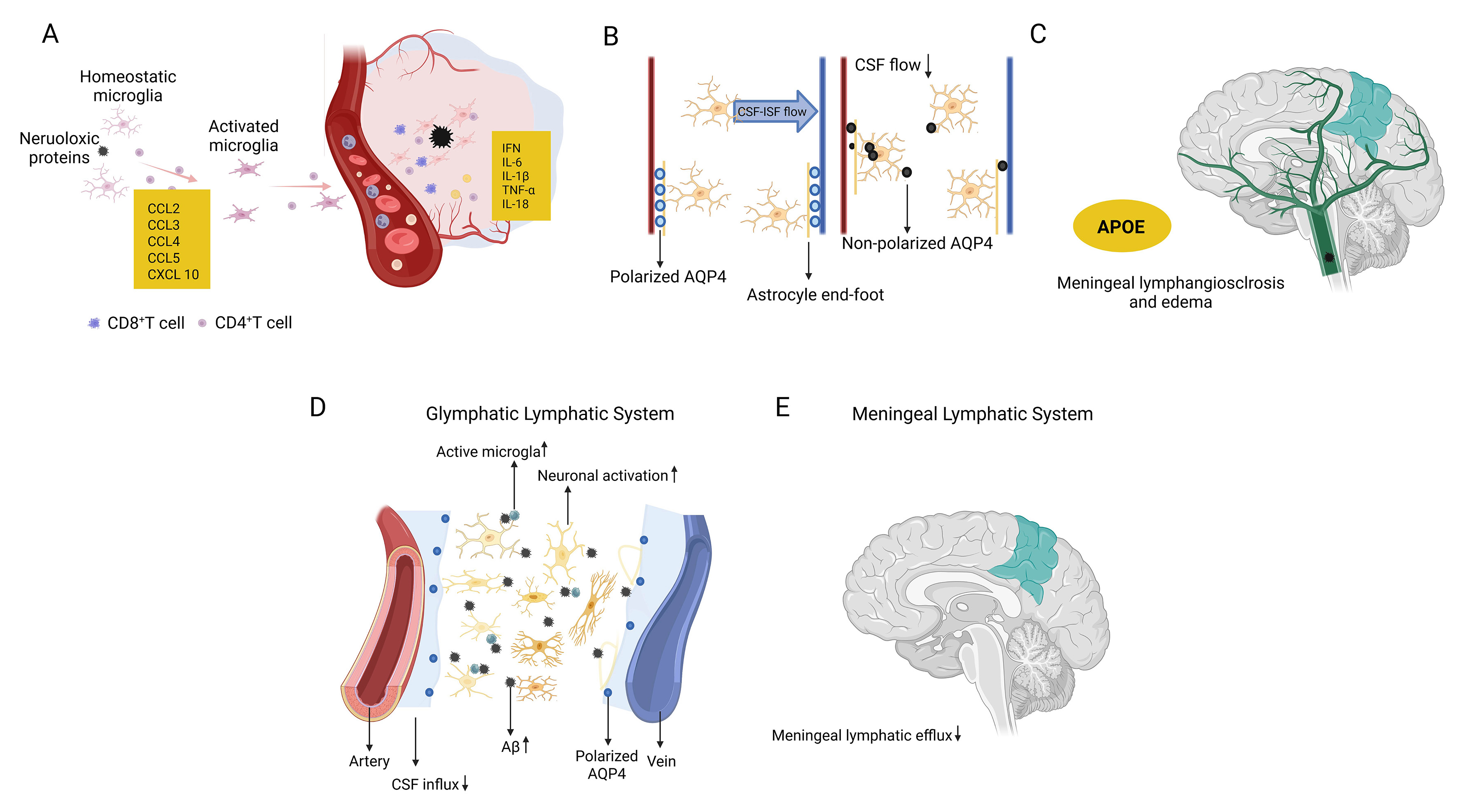
Figure 3. Glymphatic and meningeal lymphatic pathways involved in brain waste clearance in AD. (A) Homeostatic microglia become activated in response to neurotoxic protein accumulation, accompanied by the involvement of CD8+ and CD4+ T cells; (B) Aberrant loss of perivascular AQP4 polarization in astrocytes impairs CSF-ISF exchange and glymphatic flow; (C) Effects of the APOE genotype on meningeal lymphangiectasia (vessel dilation) and edema formation; (D) Schematic illustration of glymphatic and meningeal lymphatic waste clearance pathways, showing increased CSF influx associated with microglial activation and neuronal activity; (E) Illustration of the meningeal lymphatic system, including arteries, increased CSF influx, the posterior vein with AQP4, and impaired meningeal lymphatic efflux. This figure was designed by the authors using BioRender. https://BioRender.com/adw2nwf. AD: Alzheimer’s Disease; Aβ: amyloid-beta; APOE: apolipoprotein E; AQP4: aquaporin-4; CCL2: C-C motif chemokine ligand; CXCL 10: C-X-C motif chemokine ligand 10; CD8+ T cell: cluster of differentiation 8-positive T cell; CSF: cerebrospinal fluid; ISF: interstitial fluid; INF: interferon; TNF-α: tumor necrosis factor-α; IL-1β: interleukin-1β.
THERAPEUTIC STRATEGIES TARGETING GLYMPHATIC AND LYMPHATIC SYSTEMS IN AD
Current U.S. Food and Drug Administration (FDA)-approved therapies for AD, such as acetylcholinesterase inhibitors and N-methyl-D-aspartate receptor antagonists, provide only limited symptomatic benefits but fail to halt disease progression[58]. More recently, monoclonal antibodies targeting Aβ pathology, such as Aducanumab and Lecanemab, have shown potential in reducing Aβ burden. However, their long-term clinical efficacy and safety remain controversial[59]. Similarly, Sodium Oligomannate (GV-971), approved in China in 2019, has encouraging therapeutic potential in preliminary studies, although additional large-scale clinical validation is still required[60]. These limitations underscore the urgent need for therapies that target the root causes of AD, particularly impaired Aβ clearance and accumulation. Emerging evidence has identified the glymphatic system and mLVs as promising therapeutic targets. By enhancing endogenous brain waste clearance pathways, future interventions could facilitate the removal of Aβ and other neurotoxic metabolites, thereby slowing or modifying AD progression [Figure 4][23].
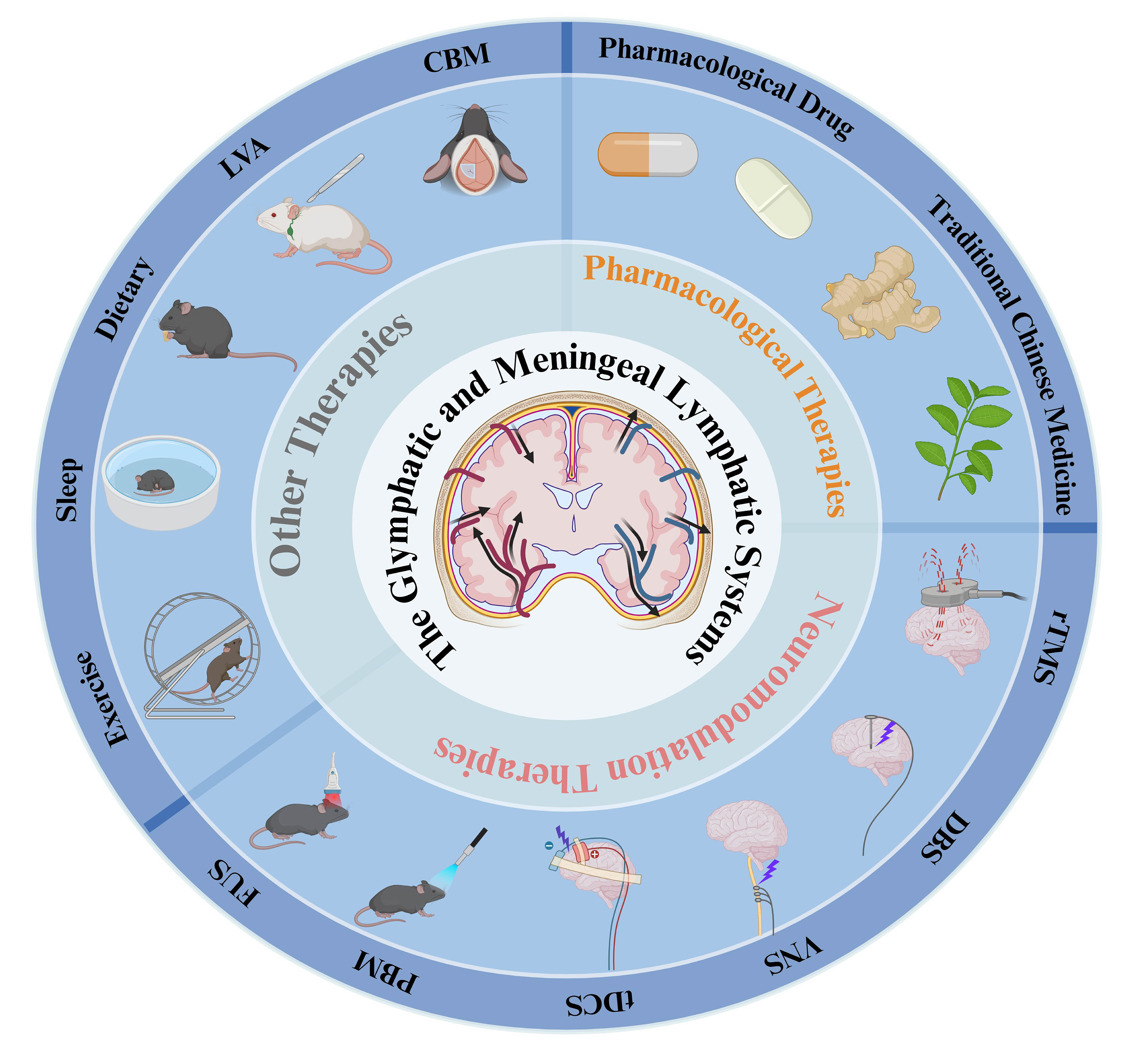
Figure 4. Multimodal therapeutic strategies targeting glymphatic and meningeal lymphatic function in AD. This schematic summarizes multiple therapeutic approaches designed to enhance brain clearance through modulation of the glymphatic and meningeal lymphatic systems. The central illustration depicts fluid dynamics (CSF/ISF flow) within these clearance pathways, whereas the surrounding concentric rings classify the interventions into pharmacological therapies, neuromodulation-based approaches, and other therapeutic strategies. These approaches augment key clearance processes, such as AQP4 polarization, CSF influx, and interstitial fluid drainage. In AD models,improved glymphatic and meningeal lymphatic function has been associated with attenuated Aβ deposition and improved cognitive impairment, highlighting the therapeutic potential of targeting brain clearance pathways in AD. This figure was designed by the authors using BioRender. https://BioRender.com/u5q9as0. AD: Alzheimer’s Disease; Aβ: amyloid-beta; AQP4: aquaporin-4; CBM: cranial bone maneuver; CSF: cerebrospinal fluid; DBS: deep brain stimulation; ISF: interstitial fluid; LVA: lymphovenous anastomosis; FUS: focused ultrasound; PBM: photobiomodulation therapy; rTMS: repetitive transcranial magnetic stimulation; tDCS: transcranial direct current stimulation; VNS: vagus nerve stimulation.
Pharmacological therapies
Pharmacological drug therapies
Age-related declines in melatonin secretion have been closely associated with AD progression, and reduced CSF melatonin levels correlate with increased pathological severity[61]. Emerging evidence indicates that melatonin is also involved in regulating glymphatic and lymphatic drainage function. Specifically, melatonin enhances glymphatic clearance and promotes the removal of Aβ aggregates. In Tg2576 transgenic mice, melatonin treatment increased soluble Aβ levels in cervical lymph nodes while reducing oligomeric Aβ accumulation in the brain, suggesting improved glymphatic-mediated waste clearance[62]. Furthermore, in patients with mild-to-moderate AD, adjunctive treatment with prolonged-release melatonin improved cognitive function and significantly alleviated sleep disturbances, highlighting its dual therapeutic roles in sleep regulation and Aβ clearance[63]. A recent study examining alterations in Aβ deposition following pharmacological modulation of AQP4 demonstrated that administration of the AQP4 facilitator N-[3-(Benzyloxy)pyridin-2-yl] benzenesulfonamide (TGN-073) significantly enhanced Aβ clearance, reduced amyloid deposition, and improved cognitive performance in mice. In contrast, treatment with the AQP4 inhibitor N-(1,3,4-thiadiazol-2-yl)pyridine-3-carboxamide dihydrochloride (TGN-020) exacerbated Aβ accumulation and behavioral deficits. Collectively, these findings indicate that modulation of AQP4 represents a promising therapeutic strategy for augmenting Aβ clearance and mitigating AD pathology[64]. Notably, melatonin may also exert neuroprotective effects through additional mechanisms, including anti-inflammatory activity and circadian rhythm regulation, both of which could independently influence Aβ and tau pathology. However, within the scope of this review, particular emphasis is placed on the role of melatonin in promoting glymphatic clearance[61-64].
Recent studies have demonstrated that glucagon-like peptide-1 receptor (GLP-1R) signaling significantly enhances Aβ clearance in AD models by modulating the subcellular localization of astrocytic AQP4. Chronic administration of the GLP-1R agonist liraglutide in AD mice activates the protein kinase A (PKA) signaling pathway and induces phosphorylation of AQP4 at Ser276, thereby reversing the depolarized distribution of AQP4 in reactive astrocytes and restoring its localization to perivascular endfoot membranes. These findings indicate that the GLP-1R-PKA-AQP4 axis constitutes a critical molecular pathway involved in Aβ clearance and reveal a novel mechanism by which GLP-1R agonists restore AQP4-mediated glymphatic function to counteract AD pathology. These findings provide a mechanistic foundation for the development of metabolism-based therapeutic strategies for neurodegenerative diseases[65].
Recent studies have revealed that Butylphthalide (NBP) exhibits potential neuroprotective effects in mouse models of AD. Short-term administration of NBP enhances the pulsatility of both superficial and penetrating cerebral arteries, thereby accelerating glymphatic clearance and subsequent lymphatic drainage, ultimately reducing parenchymal Aβ accumulation in APP/PS1 mice[66]. A preclinical study further demonstrated that the novel small-molecule compound OAB-14 enhances glymphatic function, promotes cerebral Aβ clearance, and alleviates cognitive impairment in AD mouse models by upregulating AQP4 expression and preserving its polarized localization through the PPARγ-P2X7r-AQP4 signaling pathway[67].
Oxytocin (OT), a neuropeptide with anti-inflammatory properties, has shown potential therapeutic effects in improving cognitive function in AD. Recently, OT enhances intracranial lymphatic drainage in AD mouse models by improving cerebral blood flow, inhibiting edema-associated proteins, and promoting lymphangiogenesis. These effects restored the structural integrity and drainage efficiency of the lymphatic system, thereby facilitating Aβ clearance in middle-aged APP/PS1 mice[68].
Cilostazol, a vasoactive agent, promotes Aβ clearance in AD mouse models[69] and induces the proliferation and stabilization of lymphatic endothelial cells through the activation of the PKA/p-CREB signaling pathway, thereby accelerating waste clearance[70]. Based on these findings, researchers designed a long-acting hydrophilic liquid crystal implant encapsulating cilostazol and Donepezil hydrochloride, which was administered through injection into the deep cervical lymph nodes of mice. This experimental delivery method increased lymphatic vessel coverage in the meninges and cervical lymph nodes, enhanced macromolecular drainage, and reduced Aβ deposition. Additionally, treated mice exhibited significant improvements in learning ability and cognitive performance[6].
VEGF-C is critical for the development of mLVs in the brain’s membranes[71]. In mice, VEGF-C intervention induces meningeal lymphangiogenesis and increases luminal diameter[72]. To investigate the synergistic effects of enhanced meningeal lymphatic drainage combined with anti-Aβ monoclonal antibodies on plaque clearance in AD mice, researchers combined the mAb158 antibody with VEGF-C to explore its therapeutic potential. The results showed that in 5xFAD mice, Aβ deposition and microglial activation were reduced, and lymphatic vessels surrounding the transverse sinus were significantly expanded[21].
Studies have found that a hyperosmolar plasma state significantly enhances the delivery and distribution of Aβ antibodies in the brain by reversing glymphatic drainage impairment in AD mice, thereby improving brain dysfunction[73]. Additionally, high doses of the α2-adrenergic receptor agonist dexmedetomidine can enhance CSF glymphatic influx and promote the delivery of Aβ antibodies to the brain[74]. These strategies enhance therapeutic antibody efficacy, providing a novel strategy to optimize immunotherapy for AD.
Beyond Aβ, glymphatic-targeted interventions also show promise in addressing tau pathology. Chronic inhibition of AQP4 using TGN-020 in tauopathy models reduces tau aggregation and propagation, underscoring the role of the glymphatic system in tau clearance[75]. This emerging focus on tau pathology highlights the potential of dual-target therapies for addressing the multifactorial nature of AD. However, these pharmacokinetic and pharmacodynamic considerations, together with the inherent complexity of human glymphatic physiology, warrant caution in interpreting preclinical efficacy. The stark reality of AD drug development, with failure rates exceeding 99%, underscores the limited predictive validity of current transgenic models in recapitulating human disease pathology. Furthermore, critical pharmacological parameters, including BBB penetration, optimal dosing, and target engagement, remain largely unaddressed for most of these compounds, emphasizing substantial barriers to clinical translation[21,61-65,67,74,75].
TCM therapies
TCM, grounded in holistic principles and personalized treatment strategies, offers multi-target therapeutic potential for AD. Emerging research highlights that TCM can enhance glymphatic clearance, reduce Aβ deposition, and improve cognitive function through diverse mechanisms[76].
Borneol, a natural compound, improves meningeal lymphatic drainage by enhancing vessel permeability and expanding luminal diameter in mice. This effect reduces nitric oxide levels in the meninges, decreases Aβ accumulation, and ameliorates cognitive deficits in KM mice[77]. Similarly, Yuanzhi Powder, a classical TCM formula, restores AQP4 polarization, inhibits reactive astrogliosis, and boosts interstitial waste transport, thereby facilitating Aβ clearance from CSF to deep cervical lymph nodes in APP/PS1 mice. These effects correlate with reduced cerebral Aβ burden and improved memory performance[78].
Liu et al.[79] investigated the modified Jiawei Xionghuang Decoction in AD by establishing a model of impaired meningeal lymphatic drainage through ligation of the deep cervical lymph nodes in APP/PS1 mice. Jiawei Xionghuang Decoction restores cervical lymphatic drainage in AD mice by suppressing inflammatory pathways associated with arachidonic acid metabolism, thereby enhancing Aβ clearance and alleviating cognitive impairment[79]. Xueshuantong, a therapeutic agent for stroke[80], also improves lymphatic vessel function and mitigates neuroinflammation in AD mice[81]. Additionally, ischemia-reperfusion injury following stroke can induce rapid cerebral Aβ deposition, thereby exacerbating cognitive impairment[82]. Tongxinluo, a commonly prescribed formula for cardiovascular and cerebrovascular diseases, has been reported to block inflammatory cell death (pyroptosis), restore AQP4 expression, and reduce Aβ accumulation after cerebral ischemia-reperfusion in rats[83].
The Yizhi Formula further demonstrates the versatility of TCM by restoring the interactions between the BBB, glymphatic system, and meningeal lymphatic system. It inhibits inflammatory glial activation, reduces Aβ oligomer formation, and enhances waste clearance in preclinical models[84]. Collectively, these findings underscore the ability of TCM to target AD through complementary mechanisms, including attenuation of neuroinflammation, optimization of fluid dynamics, and preservation of vascular integrity. By leveraging natural compounds to enhance glymphatic function and modulate Aβ pathology, TCM represents a promising adjunct to conventional AD therapies. Further clinical studies are warranted to validate these preclinical insights and refine TCM-based interventions [Table 1]. Moreover, current evidence is derived exclusively from preclinical studies, with no randomized controlled trials (RCTs) evaluating TCM-mediated glymphatic enhancement in AD patients. The absence of placebo-controlled, double-blind designs, coupled with insufficient pharmacokinetic and toxicological data, limits the interpretability and generalizability of these findings. Future research should prioritize rigorous quality control, systematic identification of bioactive constituents, and well-designed clinical trials to validate the efficacy and safety of TCM-based interventions[78,79,81,84].
Pharmacological therapies improve glymphatic/lymphatic drainage in AD
| Drug | Drug type | Animal model | Sample size | Impact on lymphatic drainage | Neurophysiological changes | Effect size | Cognitive change | Mechanism | Author/Year/Journal | |
| Melatonin | Chemical medicine | Tg2576 mice | N = 60-80 (total), n = 2-16 per group | ↑Soluble Aβ40 monomer levels in cervical/axillary lymph nodes; ↓Aβ oligomers in the brain | ↓Aβ40/Aβ42 oligomers in the brain | ↑lymph node monomeric Aβ40 (P = 0.017) and ↑trend for Aβ42 (P = 0.1); ↑trend brain monomeric Aβ40 (P = 0.07); ↓brain oligomeric Aβ40 (P < 0.05) | / | Inhibits Aβ aggregation, modulates immune function and antioxidant activity; promotes lymphatic clearance of Aβ monomers, reduces cerebral Aβ deposition | Pappolla et al., (2018). Current Alzheimer Research[62] | |
| TGN-073 | Chemical medicine | APP/PS1 mice (2 months) | N = 57 (total), n = 12-15 per group | ↑glymphatic clearance via AQP4 polarization; ↑ISF solute clearance | ↓Aβ deposition | ↓50%-60% total Aβ burden (P < 0.05-0.001); ↓Aβ40/Aβ42 ratio (P < 0.0001); ↓27% soluble Aβ (P = 0.031); ↓36% plaque-vessel association (P < 0.01); ↓Aβ 40/Aβ42 ratio (P < 0.01); restored memory/anxiety to WT levels. TGN-020 (AQP4 inhibitor); ↑anxiety and memory deficits (P < 0.05-0.01) | ↑Learning & memory | AQP4 modulation bidirectionally regulates intramural periarterial drainage | Manescu et al., (2025). Alzheimer’s & Dementia[64] | |
| liraglutide | Chemical medicine | AppNL-G-F/NL-G-F mice | N = 60-80 (total), n = 3-20 per group | ↑ISF-to-CSF solute clearance | ↓cortical Aβ(1-42) deposition | ↓Aβ(1-42) accumulation in cerebral cortex (P < 0.0001); ↑spontaneous alternation rate in Y-maze test (P < 0.05); ↑AQP4 localization to perivascular site (P < 0.01); ↑AQP4 serine phosphorylation (P < 0.01); ↑FITC-albumin flux from perivascular to interstitial space (P < 0.01) | ↑Learning & memory | Liraglutide activates astrocytic GLP-1R→↑cAMP/PKA signaling; PKA phosphorylates AQP4 at Ser276→relocalizes AQP4 to perivascular endfeet; Restored AQP4 polarization ↑glymphatic flux→promotes Aβ efflux | Sasaki et al., (2024). Biochemical and Biophysical Research Communications[65] | |
| L-3-n-butylphthalide | Chemical medicine | APP/PS1 mice | N = 70-100 (total), n = 6-14 per group | ↑AQP4 polarization | ↓Aβ deposition | ↓Aβ (1-42) accumulation in cerebral cortex (P < 0.0001); ↑spontaneous alternation rate in Y-maze test (P < 0.05); ↑AQP4 localization to perivascular site (P < 0.01); ↑PKA activity (P < 0.05); ↑AQP4 serine phosphorylation (P < 0.01); ↑FITC-albumin flux from perivascular to interstitial space (P < 0.01) | ↑Learning & memory | Activates β1/β2 adrenergic receptors to enhance cerebrovascular pulsation, drives glymphatic Aβ clearance; upregulates AQP4 polarization to improve glymphatic function | Zhang et al., (2021). Journal of Molecular Neuroscience[66] | |
| OAB-14 | New small molecule | APP/PS1 mice (6-8 months) | ↑CSF inflow; ↑ISF solute clearance | ↓Aβ deposition; ↓neuroinflammation; ↓neuronal damage | ↑CSF tracer influx into brain parenchyma (P < 0.05-0.01); ↑CSF tracer distribution in dcLNs (P < 0.05-0.01); ↓Remaining tracer in brain parenchyma after intraparenchymal injection (P < 0.05-0.01) | ↑Learning & memory | OAB-14 activates the Nrf2/HO-1 signaling pathway, reducing oxidative stress and restoring astrocytic AQP4 polarization. This dual action enhances glymphatic influx (CSF entry) and efflux (ISF clearance), accelerating Aβ removal and suppressing neuroinflammation | Zhang et al., (2023). Neurochemistry International[67] | ||
| Oxytocin | Chemical medicine | middle-old-aged APP/PS1 mice | N = 150-200 (total), n = 3-15 per group | ↑CSF drainage into DCLNs; ↑meningeal lymphatic vessel generation | ↓Aβ deposition; ↓neuroinflammation | ↑40%-60% lymphatic drainage to dCLNs (P < 0.01); ↑25%-50% CBF and RBC velocity (P < 0.05-0.01); ↓50%-60% AQP4 depolarization (M1/M23 ratio, P < 0.01); ↓40%-60% Aβ burden in brain (P < 0.01); ↑30%-40% CSF Aβ (P < 0.05); ↑20%-50% cognitive performance (OF, Y-maze, NOR, WM, P < 0.01-0.001) | ↑Learning & memory | Enhances cerebrovascular pulsation, inhibits AQP4 depolarization, promotes VEGF-C-mediated lymphangiogenesis; regulates meningeal immune pathways to restore intracranial lymphatic structure and Aβ clearance | Ye et al., (2024). Theranostics[68] | |
| Cilostazol and donepezil hydrochloride | Chemical medicine | C57BL/6 mice | N = 80-120 (total), n = 3-18 per group | ↑LYVE-1 coverage in meningeal lymphatic vessels/DCLNs; ↑AQP4 polarization | ↓Aβ deposition; ↓neuroinflammation | ↑LYVE-1+ lymphatic coverage in dCLNs and meninges (P < 0.05-0.01); ↑Macromolecule clearance (Cy5-OVA, FITC-Aβ) from brain to dCLNs (P < 0.01-0.001);↓Aβ deposition in hippocampus and cortex (P < 0.05-0.01) | ↑Learning & memory | Activates cAMP/PKA/p-CREB pathway to promote lymphangiogenesis; donepezil inhibits AChE, synergistically reduces Aβ deposition and improves synaptic plasticity | Shan et al., (2024). ACS Nano[6] | |
| Dexmedetomidine | Chemical medicine | C57BL/6 mice | N = 80-120 (total), n = 4-7 per group | ↑CSF inflow | / | ↑CNS-to-plasma concentration ratios in multiple brain regions (P < 0.05-0.01); ↑Amyloid-β antibody delivery to brain surfaces vs. vehicle: ~2× increase (P < 0.001) | / | Reduces norepinephrine release, increases interstitial fluid volume, lowers fluid flow resistance, and enhances CSF inflow | Lilius et al., (2019). Journal of Controlled Release[74] | |
| TGN-020 | Chemical medicine | P301S mice | N = 60-80 (total), n = 3-8 per group | ↓CSF-interstitial fluid exchange; ↓AQP4 function | ↑Tau aggregation/spread; ↑neuroinflammation | TGN-020 (AQP4 inhibitor) ↓50%-93% glymphatic function (P < 0.01-0.001); ↑30%-50% tau pathology (AT8 intensity) in hippocampus (P < 0.05-0.01); ↑20%-25% transhemispheric tau propagation (P < 0.05); ↓20%-25% NOR preference index at 4 weeks (P < 0.05); ↑2-3× hindlimb clasping score at 10 weeks (P < 0.01) | ↑Learning & memory | Inhibits AQP4-mediated glymphatic function, leading to tau retention and pathological propagation; exacerbates neurodegeneration | Lopes et al., (2024). Alzheimer’s Research Therapy[75] | |
| Borneol | Chinese herbal medicine | SD rats/Kunming mice | N = 80-100 (total), n = 3-6 per group | ↑Lymphatic vessel permeability/diameter; ↑macromolecule drainage into DCLNs | ↓Aβ deposition; ↓neuronal damage | ↑50%-100% tracer accumulation in dCLNs (P < 0.01-0.001); ↑1.5-2× lymphatic drainage (ICG-NPs AUC, P < 0.01-0.001); ↑2-2.5× lymphatic vessel diameter and LYVE-1+ area (P < 0.05-0.001); ↓40%-50% meningeal NO (P < 0.001); ↓35% brain Aβ (P < 0.01); ↑54% dCLNs Aβ (P < 0.01); ↑30%-50% cognitive performance in MWM (P < 0.05-0.01) | ↑Learning & memory | Improves lymphatic permeability/diameter, reduces nitric oxide levels in meninges, stimulates lymphatic contraction, upregulates lymphangiogenic proteins to enhance macromolecule drainage | Wu et al., (2023). Theranostics[77] | |
| Yuanzhi Powder | Chinese herbal medicine | APP/PS1 mice | N = 105-150 (total), n = 3-14 per group | ↑CSF inflow; ↑AQP4 polarization | ↓Aβ deposition; ↓neuroinflammation; ↓neuronal damage | ↑TR-d3 tracer distribution in forebrain parenchyma (P < 0.05); ↑Lymphatic vessel diameter in TS (P < 0.05); ↓Aβ plaque area in hippocampus and cortex (IHC) (P < 0.01) | ↑Learning & memory | Restores AQP4-mediated glymphatic drainage and meningeal lymphatic transport, promotes Aβ clearance from brain interstitium to periphery | Li et al., (2024). Journal of Ethnopharmacology[78] | |
| Jiawei Xionggui Decoction | Chinese herbal medicine | APP/PS1 mice | N = 80-120 (total), n = 5-8 per group | / | ↓Aβ deposition; ↓neuroinflammation | ↓Aβ plaque area in hippocampus and cortex (IHC) (P < 0.05); ↑PSA in Y-maze test (P < 0.05 vs. APP/PS1-L); ↑Lyve-1+ fluorescence intensity in dural regions (P < 0.05) | ↑Learning & memory | Inhibits arachidonic acid metabolism (↓COX-2, LTD4, PGE2), reduces neuroinflammation, restores cerebral blood flow and MLVs drainage efficiency, promotes Aβ clearance and cognition | Liu et al., (2024). Phytomedicine[79] | |
| Xueshuantong | Chinese herbal medicine | APP/PS1 mice | N = 104 (total), n = 3-9 per group | ↑AQP4 polarization; ↑meningeal lymphatic vessel morphology | ↓Aβ deposition; ↓neuroinflammation; ↓neuronal damage | ↑10%-25% lymphatic vessel diameter/area (P < 0.05-0.01); ↓30%-50% Aβ in meninges/dCLNs (P < 0.01); ↓25%-35% microglial activation at 15 days (P < 0.05); ↑40%-50% GLT-1 (P < 0.01); ↓40%-50% MMP-9 (P < 0.01); ↓20%-30% AQP4 polarity (P < 0.01) | ↑Learning & memory | Modulates lymphatic structure, potentially by reducing MMP-9 and increasing GLT-1 levels to improve lymphatic drainage and cognitive function | Zheng et al., (2021). Frontiers in Pharmacology[81] | |
| Tongxinluo | Chinese herbal medicine | SD rats | N = 35 (total), n = 10 per group | ↑AQP4 polarization | ↓Aβ deposition; ↓neuroinflammation | ↓30%-50% neurological deficit scores (P < 0.05); ↓40%-60% caspase-11/GSDMD pathway proteins (P < 0.01); ↓30%-50% NLRP3, caspase-1, IL-1β, IL-6 (P < 0.01-0.05); restored AQP-4 polarity; ↓30%-40% Aβ accumulation and Aβ1-42 oligomers (P < 0.05-0.01) | / | Inhibits caspase-11/1 activation and GSDMD cleavage, reduces astrocyte pyroptosis, restores AQP4 polarization, mitigates Aβ accumulation, alleviates neuroinflammation and BBB damage | Wang et al., (2021). Evidence-Based Complementary and Alternative Medicine[83] | |
| Yi-Zhi-Fang-Dai formula | Chinese herbal medicine | SD rats | N = 85 (total), n = 8-15 per group | ↑AQP4 polarization | ↓Aβ deposition; ↓neuroinflammation; ↓neuronal damage | ↓35%-50% neurological deficit scores (P < 0.01); ↑30%-40% BBB integrity (SMI 71) (P < 0.05); ↑20%-30% AQP-4 density (P < 0.01); ↓40%-50% Aβ accumulation and Aβ 1-42 oligomers (P < 0.01); ↓40%-50% neuronal damage (P < 0.01); ↑30%-40% NeuN (P < 0.01) | / | Protects BBB integrity and AQP4 polarization, promotes Aβ clearance and blocks toxic oligomerization, mitigates neuroinflammation | Lyu et al., (2021). Frontiers in Pharmacology[84] | |
Neuromodulation therapies
Neuromodulation therapies, ranging from ultrasound and magnetic stimulation to acupuncture and photobiomodulation, represent promising strategies to enhance glymphatic and meningeal lymphatic drainage in AD. By modulating fluid dynamics, neuroinflammation, and cellular metabolism, these interventions target multiple dimensions of AD pathology.
Focused ultrasound (FUS) is a non-invasive therapeutic modality that utilizes ultrasonic energy to precisely target specific brain regions, achieving therapeutic effects via thermal or mechanical mechanisms[85]. In vitro experiments have demonstrated that 400 kHz FUS with a pulse duration of 75 ms maximally enhances solute and fluid transport in porous materials mimicking brain tissue. Preclinical studies show that low-frequency FUS enhances fluid and solute transport in brain tissue[86], improves Aβ and tau clearance, and boosts lymphatic drainage in AD mice[87]. By transiently disrupting the BBB, FUS combined with microbubble technology increases CSF efflux and venous permeability, thereby facilitating brain waste clearance[88].
Repetitive transcranial magnetic stimulation (rTMS) is a non-invasive neuromodulation technique being investigated for its potential to enhance brain waste clearance processes in AD. Researchers have investigated high-frequency rTMS in an AD mouse model to evaluate its effects on cognitive function and disease-related neuropathological changes. The study utilized 4-5-month-old 5xFAD mice, which exhibit AD-like symptoms, including memory deficits and Aβ plaque accumulation. Findings revealed that rTMS preserved recognition memory and spatial memory for novel objects and their locations over time. Importantly, rTMS significantly enhanced the efficiency of brain clearance pathways, including the glymphatic system within the brain parenchyma and mLVs. Significant reductions in Aβ deposition, along with suppression of microglial and astrocyte activation and elevated cellular proto-oncogene Fos (c-FOS) expression, were observed in the prefrontal cortex and hippocampus of rTMS-treated 5xFAD mice[10]. Besides, our team found that cerebellar intermittent theta-burst stimulation (iTBS) enhances glymphatic clearance in AD patients, as evidenced by an increased diffusion tensor image analysis along the perivascular space (DTI-ALPS) index, and is associated with improved cognitive performance[89].
Electroacupuncture (EA) is a non-pharmacological intervention that combines electrical stimulation with traditional acupuncture and is commonly used in clinical practice[9]. Accumulating evidence suggests that EA improves cognitive performance, alleviate neuropsychiatric symptoms, and enhances daily functioning in patients with dementia[90,91]. A study in 7-month-old SAMP8 mice revealed that EA enhances lymphatic drainage and improves the structural organization of AQP4-related cellular functions[92]. Similarly, another study found that EA attenuates inflammatory responses, suppresses the activation of astrocytes and microglia, and upregulates the expression of specific hippocampus markers in APP/PS1 transgenic mice. Additionally, EA promotes lymphatic system activity[93].
Photobiomodulation therapy (PBM) is a non-invasive treatment that utilizes specific wavelengths of low-intensity light to stimulate cellular activity[94]. PBM primarily utilizes light within 400-1,300 nm range to activate mitochondrial cytochrome c oxidase, triggering a photobiochemical cascade involving Nitric Oxide (NO) release, enhanced adenosine triphosphate (ATP) production, reactive oxygen species (ROS) modulation, and activation of redox-sensitive signaling pathways that ultimately regulate cellular metabolism and function through gene expression changes[95-98]. PBM has been widely explored for the treatment of neurodegenerative diseases[99]. A pilot study demonstrated that transcranial PBM (tPBM) at 1,267 nm significantly reduces Aβ plaque burden, improves cognitive function in AD mice, and enhances brain clearance via the lymphatic system, suggesting its potential as a non-pharmacological therapeutic strategy for AD[100]. One study demonstrated that a 9-day course of 1,267 nm tPBM (32 J/cm2) significantly improves neurocognitive function (P < 0.05) and cerebral oxygenation in Aβ-injected AD mice by enhancing meningeal lymphatic clearance of Aβ and alleviating cerebral hypoxia (72% ± 6% to 97% ± 2%, P < 0.001)[101]. Another study found that PBM enhances mitochondrial metabolism and intercellular connectivity in meningeal lymphatic endothelial cells, which may underlie its beneficial effects on glymphatic drainage[102]. Low-intensity 40 Hz blue light has showed therapeutic potential in 5xFAD mice by restoring astrocytic AQP4 polarity, improving brain drainage efficiency, and reducing hippocampal lipid accumulation. When combined with anti-Aβ antibodies, it further enhances Aβ clearance and improves cognitive function[103].
Deep brain stimulation (DBS) involves the delivery of electrical pulses to specific brain regions, such as the subthalamic nucleus or globus pallidus, via implanted electrodes. It’s used to treat conditions such as Parkinson’s disease, essential tremor, and obsessive-compulsive disorder[104]. DBS has been shown to promote neuroprotection, reduce inflammation, and potentially increase cerebral blood flow and stimulate lymphatic system activity[105-107]. However, whether these changes translate into improved CSF-ISF exchange or Aβ clearance has not been demonstrated. Thus, the proposed link between DBS and glymphatic enhancement remains speculative and requires systematic investigation in dedicated preclinical models[105-107].
Vagus nerve stimulation (VNS) is an FDA-approved neuromodulation therapy used to treat epilepsy, depression, and cluster headaches[108]. An animal study demonstrated that VNS may enhance glymphatic drainage by increasing CSF penetration into the brain, thereby potentially modulating CSF/ISF exchange[109].
Transcranial direct current stimulation (tDCS) delivers a weak direct current to the scalp to modulate cortical neuronal excitability, thereby influencing brain function[110]. An animal study found that tDCS enhances glymphatic influx and efflux through astrocytic IP3/Ca2+ signaling and modulation of brain oscillatory activity, thereby promoting brain metabolic waste clearance[111]. Although the studies summarized below demonstrate promising effects, many of these modalities are supported by a limited number of preclinical investigations, often with small sample sizes and incomplete mechanistic characterization. Furthermore, optimal stimulation parameters (e.g., frequency, intensity, duration) for glymphatic enhancement have not been systematically established, and substantial heterogeneity in experimental protocols, together with a lack of standardization, complicates cross-study comparisons and impedes clinical translation. Critically, long-term safety and efficacy in humans remain largely unexplored. Future research should prioritize independent replication, rigorous dose-response analyses, and mechanistic studies leveraging advanced imaging and genetic tools to address these gaps[10,92,100,101,109,111][Table 2].
Neuromodulation therapies enhance glymphatic drainage function in AD
| Neuromodulation therapies | Animal model | Sample size | Impact on lymphatic drainage | Neurophysiological changes | Effect size | Cognitive behavioral changes | Mechanism of action | Author/Year/Journal |
| PBM | ||||||||
| tPBM | C57 mice | N = 140 (total), n = 7 per group | ↑CSF drainage into DCLNs | ↓Aβ deposition | Cohen’s d ≈ 2.5-3.0 for Aβ reduction; Cohen’s d ≈ 3.0-4.5 for increased lymphatic clearance | ↑Learning and memory | Not investigated | Zinchenko et al. (2019). Biomedical Optics Express[100] |
| tPBM | C57 mice | N = 40 (total), n = 10 per group | ↑CSF drainage into DCLNs | ↓Aβ deposition | ↓Aβ plaques (P < 0.001); ↑GNR accumulation in dCLNs (AAS, P < 0.001); ↑SpO₂ (72%→85%, P < 0.001); ↓NSS (3.67→2.03, P < 0.05) | ↑Learning and memory | Improved oxygen consumption and enhanced mitochondrial production of adenosine triphosphate | Semyachkina-Glushkovskaya et al. (2021). Advances in Experimental Medicine and Biology[101] |
| near-infrared light | (5xFAD and APP/PS1) mice | N = 150-200 (total), n = 5-21 per group | ↑CSF infux | ↓Aβ deposition; ↓neuroinflammation; ↓neuronal damage | ↑lymphatic drainage (P < 0.001); ↑cognitive performance (NOR, P < 0.001); ↓Aβ deposition (P < 0.001); ↑mitochondrial length in mLECs (P < 0.001) | ↑Learning and memory | Improving mitochondrial metabolic function of meningeal lymphatic endothelial cells | Wang et al. (2024). Nature Communications[102] |
| 40HZ blue light | 5xFAD mice | N = 150-200 (total), n = 6-21 per group | ↑CSF drainage into DCLNs | ↓Aβ deposition; ↓neuroinflammation | ↑cognitive performance (Y-maze, NOR, P < 0.05-0.001); ↑glymphatic drainage (dCLNs, P < 0.01-0.001); ↑AQP4 polarity (P < 0.05-0.001); ↓soluble Aβ42 (P < 0.05). | ↑Learning and memory | Activation of the vLGN/IGL-Re-mediated visual pathway restores AQP4 polarity | Wu et al., (2025). Nature Communications[103] |
| TMS | 5xFAD mice | N = 80-100 (total), n = 7-13 per group | ↑CSF infux | ↓Aβ deposition; ↓neuroinflammation; ↓neuronal damage | ↑cognitive performance (NOR/NOL, P < 0.05-0.01); ↓Aβ deposits (P < 0.05-0.001); ↑glymphatic/meningeal lymphatic drainage (P < 0.05-0.01); ↓glial activation (P < 0.05-0.001); ↑neuronal activity (c-FOS, P < 0.05-0.001). | ↑Learning and memory | Not investigated | Lin et al., (2021). Acta Neuropathologica Communications[10] |
| TMS | AD patients | 20 patients | ↑Diffusion tensor image analysis along the perivascular space | Not investigated | Change in CDR-SB score from baseline to week 4: Active-iTBS group: -0.60. Sham-iTBS group: +0.15. Adjusted β = 0.73 (95%CI: 0.17-1.26) | ↑Memory | Not investigated | Zhang et al. (2025). Journal of Alzheimer’s Disease[89] |
| FUS | 5xFAD mice | N = 60-80 (total), n = 6-10 per group | ↑CSF drainage into DCLNs | ↓Aβ deposition; ↓neuroinflammation; ↓neuronal damage | ↓ Aβ deposits (IHC, P < 0.05-0.005); ↑CSF Aβ1-42 after dCLN ligation + FUS-MB (P < 0.005); ↑working memory in ligated mice (Y-maze, P < 0.05); ↑meningeal vascular amyloid accumulation (P < 0.05) | ↑Learning and memory | Not investigated | Lee et al., (2020). Scientific Reports[87] |
| FUS | / | N = 8 (total), n = 3 per group | ↑CSF infux | Not investigated | 100% success in BBB opening and perivenous enhancement; 100% venous permeabilization (24-48 h); 100% CSF effusions (size ↑1.2-5.3 cm) | Not investigated | Not investigated | Mehta et al., (2023). Fluids and Barriers of the CNS[88] |
| EA | SAMP8 mice | N = 45 (total), n = 3-6 per group | ↑CSF infux | ↓Aβ deposition; ↓neuronal damage | ↑cognitive performance (MWM, P < 0.05-0.001); ↑neuronal survival (Nissl, P < 0.05); ↓Aβ1-40/1-42 (ELISA, P < 0.05-0.01); ↓Gd-DTPA retention (MRI, P < 0.05-0.01); ↓astrocyte reactivity (GFAP, P < 0.05-0.01); ↑AQP4 polarity (P < 0.05) | ↑Learning and memory | Restore AQP4 polarity | Liang et al., (2021). Neural Plasticity[92] |
| EA | C57BL/6 mice | N = 40 (total), n = 10 per group | Not investigated | ↓Aβ deposition; ↓neuronal damage | ↑preference index (P < 0.05); ↓Aβ1-40/1-42 (P < 0.01-0.05); ↓GFAP, Iba-1 (P < 0.01-0.05); ↓IL-1β, IL-6, TNF-α (P < 0.01-0.05); ↑AQP4 polarity (P < 0.05) | ↑Learning and memory | Restore AQP4 polarity | Wang et al., (2020). Zhen Ci Yan Jiu[93] |
Exercise enhances glymphatic and lymphatic drainage in AD
Physical exercise is a well-established non-pharmacological strategy for the prevention and management of neurological disorders, including AD[112,113]. It is widely recognized as one of the most effective methods to enhance cognitive function[114]. Clinical meta-analyses consistently demonstrate that higher levels of physical activity are associated with reduced AD incidence and slowed disease progression[115]. Preclinical studies further suggest that exercise enhances the clearance of pathological proteins such as Aβ, potentially by improving glymphatic system function, a key pathway for brain waste removal[13,116].
Animal studies provide additional mechanistic insights into these effects. For example, studies in aged mice demonstrated that voluntary exercise enhances astrocytic AQP4 expression and polarization without affecting BBB integrity. These changes correlated with reduced inflammation, fewer Aβ plaque burden, and strengthened glymphatic drainage in the brain[117]. Similar findings were observed in young mice following five weeks of voluntary wheel running, where exercise increased CSF movement and waste clearance efficiency across brain regions, suggesting a direct association with improved cognitive performance[118]. A recent study demonstrated that aerobic exercise training enhances recovery of learning and memory function in an AD mouse model by modulating glymphatic system activity[119].
Accumulating evidence demonstrates that polarized distribution of astrocytic AQP4 is critical for exercise-induced enhancement of glymphatic function. Liu et al.[120] revealed that AQP4-knockout APP/PS1 mice exhibit exacerbated impairments in glymphatic function, increased Aβ deposition, and more severe cognitive deficits, which could not be ameliorated by exercise intervention. Besides, genetic ablation of AQP4 in AD experimental models may abolish exercise-induced improvements in spatial learning and memory[120]. Conversely, recent studies show that exercise in aged mice restores physiological AQP4 polarization by modulating TRPV4-mediated calcium signaling in brain cells, thereby reducing amyloid accumulation and protecting against cognitive decline[121].
High-Intensity Interval Training (HIIT), an efficient and time-saving exercise modality, has been reported to enhance hippocampal-dependent memory and neurogenesis in rodents[85]. Notably, HIIT has also been shown to ameliorate cognitive deficits in AD[122]. A foundational study demonstrated that 8 weeks of HIIT promotes redistribution (depolarization) of phosphorylated and non-phosphorylated AQP4 in the cerebral cortex and hippocampus of streptozotocin-induced AD rat models. Furthermore, HIIT modulates AQP4 polarization by facilitating astrocytic phenotypic transition from the neurotoxic A1 state to the neuroprotective A2 state. Restored AQP4 polarization enhances the clearance of Aβ and tau proteins via the glymphatic system and renal pathways, ultimately improving cognitive deficits and neuronal damage in AD rats[123]. These findings highlight the dual role of exercise in supporting brain health through both structural and molecular mechanisms.
Sleep modulation improves glymphatic drainage in AD
Sleep disturbances are strongly linked to AD progression, with population studies revealing a direct relationship between disrupted sleep patterns and accelerated cognitive decline[124]. More than half of patients with AD experience sleep disturbances, making sleep disorders a significant risk factor for disease development and progression[125]. Emerging evidence underscores the critical role of sleep in maintaining brain homeostasis through activation of the glymphatic system, which is essential for the clearance of neurotoxic proteins such as Aβ and tau[18]. Disruption of circadian rhythms directly impairs γ-secretase activity, which itself exhibits circadian oscillations, leading to aberrant processing of amyloid precursor protein (APP) and increased production and accumulation of Aβ peptides. Concurrently, Aβ accumulation further disrupts sleep-wake regulation, establishing a bidirectional and self-reinforcing pathological feedback loop[126]. Accordingly, maintaining healthy sleep habits is crucial for brain health and may play a key role in preventing AD.
Ju et al. reported that disruptions in slow-wave sleep (SWS) increase CSF Aβ levels, whereas subacute sleep disturbances are associated with elevated CSF tau levels[11]. This is likely because the glymphatic system, responsible for brain waste clearance, is most active during sleep[18]. Sleep deprivation significantly impairs glymphatic system function[28]. Both clinical and preclinical studies indicate that sleep regulates levels of neurotoxic AD-related proteins in the brain. The sleep-wake cycle modulates Aβ levels in both ISF and CSF, whereas chronic sleep deprivation exacerbates tau accumulation and promotes tau propagation[32].
Notably, enhancing slow-wave activity (SWA) during non-rapid eye movement (NREM) sleep has emerged as a potential therapeutic strategy. By enhancing glymphatic clearance, interventions targeting sleep quality, such as optimizing sleep duration or modulating specific sleep stages, may reduce Aβ deposition and slow neurodegeneration[127]. Although sleep modulation influences multiple physiological processes, including synaptic homeostasis, hormone secretion, and inflammation, its effects on glymphatic function represent a key mechanism underlying reduced Aβ and tau accumulation. This review focuses on the glymphatic-mediated effects of sleep, while acknowledging that additional pathways may also contribute to its therapeutic potential[11,32].
Other methods enhance glymphatic drainage in AD
Epidemiological evidence suggests that lifestyle and dietary factors significantly influence AD risk and disease progression[128]. Nutritional supplementation, particularly high-dose n-3 polyunsaturated fatty acids (PUFAs), has shown promise in preclinical studies[129]. In fat-1 transgenic mice, PUFAs significantly enhance interstitial Aβ clearance in the brain. Notably, the same intervention fails to produce effects in AQP4-knockout mice, further validating that n-3 PUFAs modulate glymphatic function by suppressing astrocyte activation and preserving AQP4 polarization in affected brain regions, thereby reducing Aβ deposition[130]. Meanwhile, a recent animal study demonstrated that high-dose fish oil supplementation significantly reduced Aβ accumulation in retinal vessels. This effect is potentially mediated by regulation of key proteins involved in transcytosis, including Mfsd2a, as well as aquaporin AQP4, suggesting its potential as an adjunct therapeutic strategy for AD-related retinopathy[131]. Conversely, long-term high-fat diet, a known risk factor for AD, impairs glymphatic system function by disrupting the polarization of AQP4. This results in reduced clearance and accelerated Aβ deposition in the brain[17].
Cervical deep lymphovenous anastomosis (LVA) has emerged as a novel surgical approach for moderate-to-severe AD. This procedure involves anastomosing bilateral deep cervical lymphatic vessels to veins, thereby redirecting glymphatic and meningeal lymphatic outflow from the brain to deep cervical veins for extracranial drainage. By decompressing the lymphatic outflow pathway, LVA facilitates lymphatic fluid drainage from high-pressure lymphatic vessels into the low-pressure venous system, enhancing CSF dynamics and promoting clearance of AD-related toxic proteins, including Aβ and tau[132]. A clinical case report showed that, five weeks postoperatively, patients exhibited improved cognitive function and alleviation of mood symptoms, such as depression. Besides, tau-positron emission tomography (PET) imaging revealed reduced global brain tau accumulation[133]. A recent preclinical study established the first rat model of deep cervical lymph node-venous anastomosis (LNVA) using high-magnification microsurgical techniques to perform bilateral cervical LNVA, providing a standardized platform for investigating glymphatic system function and its regulatory mechanisms[134]. A single-arm exploratory study demonstrated that deep cervical LVA is a safe and feasible procedure for patients with AD, significantly improving cognitive function as measured by the Mini-Mental State Examination (MMSE) at one month postoperatively, although other neuropsychological outcomes and CSF biomarkers did not reach statistical significance[135]. More recently, a prospective single-arm study evaluated the therapeutic potential of deep cervical lymphatic-venous anastomosis in patients with severe AD. This intervention was associated with slowed cognitive decline and improvements in core biomarker profiles (e.g., CSF Aβ42, p-tau 181), suggesting that surgical enhancement of lymphatic drainage may directly facilitate clearance of neurotoxic proteins in humans[136].
Another innovative approach involves mechanical modulation through cranial bone maneuver (CBM) in mouse models of AD. This technique stimulates de novo mLV formation at key sinus regions (superior sagittal and transverse sinuses) and enhances lymphatic clearance. It also primes the skull-meninges interface for inflammatory activation and subsequent lymphangiogenesis. Sustained improvements in cognitive deficits and amyloid pathology at 60-day follow-up demonstrate the long-lasting neuroprotective potential of CBM. These approaches highlight the therapeutic potential of modulating brain fluid dynamics through structural or mechanical interventions [Table 3].
Other Therapies to Improve Glymphatic/Lymphatic Drainage in AD
| Therapies | Intervention method | Animal model | Sample size | Impact on lymphatic drainage | Neurophysiological changes | Effect size | Cognitive behavioral changes | Mechanism of action | Author/Year/Journal |
| Exercise | Voluntary wheel running exercise | Aged mice | N = 36 (total), n = 6 per group | ↑ISF solute clearance rate; ↑CSF drainage into dCLNs | ↓Aβ; ↓neuroinflammation; ↓neuronal damage | ↑40%-50% cognitive performance (MWM, P < 0.001); ↑1.5- to 2-fold glymphatic clearance (tracer in dCLNs, P < 0.01); ↑20%-86% AQP4 expression/polarity (P < 0.05-0.001); ↓35%-44% glial activation (GFAP, Iba1, P < 0.05-0.001); ↓49%-70% Aβ accumulation (ELISA/IHC, P < 0.01-0.001) | ↑Learning & memory | Enhances AQP4 expression and polarization at perivascular endfeet; increases dendritic spine density | He et al. (2017). Frontiers in Molecular Neuroscience[117] |
| Exercise | Voluntary wheel running | Young female C57BL/6J mice | N = 38 (total), n = 3-6 per group | Awake mice: ↑CSF inflow; Acute exercise: ↓CSF inflow inhibition; No effect under anesthesia | No change in GFAP/AQP4 expression; AQP4 polarization trend | ↑2-fold glymphatic influx in awake exercised mice (P < 0.05); ↑tracer in hypothalamus, ventral/lateral cortex (P < 0.01-0.001); ↓10% heart rate (P = 0.006); ↓50%-60% tracer influx during acute running (P < 0.05) | / | Long-term exercise enhances lymphatic activity via cardiovascular regulation and reduced noradrenergic inhibition; acute exercise inhibits due to increased norepinephrine signaling | von Holstein-Rathlou et al. (2018). Neuroscience Letters[118] |
| Exercise | Aerobic exercise training | Male APP/PS1 mice (4 months) | N = 80-100 (total), n = 3-10 per group | ↑CSF-ISF exchange efficiency | ↓Hippocampal Aβ plaques; ↓neuroinflammation | ↓Escape latency in Barnes maze (days 2-5) (P < 0.001); ↓Aβ plaque area fraction in hippocampus (IHC) (P < 0.001); ↑TR-d3 tracer diffusion area in brain parenchyma (P < 0.01-0.001) | ↑Learning & memory | Swimming↑Lama1/Dp71 expression→stabilizes AQP4 polarization at perivascular endfeet; Enhanced AQP4 water flux→↑CSF-ISF exchange→promotes Aβclearance. | Liang et al. (2025). Brain Research Bulletin[119] |
| Exercise | Voluntary wheel exercise | APP/PS1 mice (3 months) | N = 144-156 (total), n = 4-13 per group | ↑CSF inflow | ↑AQP4 polarization; ↑synaptic proteins; ↓Aβ; ↓neuroinflammation | Large effect in 5-month APP/PS1 mice: ↑20%-40% cognitive performance (MWM, Y-maze, NOR, P < 0.05-0.001); ↑40%-50% glymphatic influx (P < 0.05-0.01); ↓30%-60% Aβ deposition (IHC, ELISA, P < 0.05-0.001); ↓20%-35% glial activation and neuroinflammation (P < 0.05-0.01); ↑30%-40% AQP4 polarity (P < 0.05); ↑30%-60% BDNF signaling and synaptic proteins (P < 0.05-0.001) | ↑Learning & memory | Improves AQP4 polarization | Liu et al. (2022). Neural Regeneration Research[120] |
| Exercise | High-intensity interval training | APP/PS1 mice (7 months) | N = 112-128 (total), n = 6-16 per group | No significant change in CSF inflow | No change in AQP4 polarization, synaptic proteins, Aβ, or neuroinflammation | ↑30%-50% cognitive performance ( P < 0.05); ↓45%-60% neuronal degeneration/apoptosis (P < 0.05); ↑25%-45% synaptic/structural proteins (MAP2, MBP, synaptophysin, spinophilin, P < 0.05); ↓25%-40% glial activation (GFAP, Iba-1, P < 0.05); ↑40%-60% A2/M2 phenotype (S100A10, CD206, P < 0.05); ↓35%-50% A1/M1 phenotype (C3d, CD86, P < 0.05); ↑35%-50% AQP4 polarity (P-AQP4, n-AQP4, P < 0.05); ↓35%-50% brain Aβ/p-tau (P < 0.05) | No significant cognitive change | AQP4 deficiency | Feng et al. (2023). Theranostics[123] |
| Exercise | Voluntary wheel exercise | Old C57BL/6J female mice | N = 80-100 (total), n = 3-11 per group | / | ↑AQP4 polarization; ↓Aβ; ↓neuroinflammation | ↑CSF tracer (FITC-dextran) influx and clearance (P < 0.01); ↓Aβ1-40 fluorescence in cortex and hippocampus (P < 0.01); ↓Aβ1-42 fluorescence in cortex and hippocampus (P < 0.01-0.05) | ↓Cognitive decline & anxiety-like behavior | Inhibits TRPV4 expression and TRPV4-associated calcium hyperactivity, promoting AQP4 polarization and modulating astrocyte phenotypes | Li et al. (2024) Experimental Neurology[121] |
| Sleep Modulation | Sleep deprivation | SD mice | N = 80-100 mice (total) + 6 human participants, n = 4-16 per group | ↓ISF-CSF convective exchange | ↑Aβ & tau; ↑neuroinflammation | ↑90%-100% mouse ISF tau during wakefulness/SD (P < 0.01); ↑51.5% human CSF tau during SD (P < 0.001); ↑36.4% CSF α-synuclein (P < 0.01); ↑50%-70% tau spreading to LC (P < 0.01); ↑30% ISF Aβ and ↑40% ISF tau via chemogenetic wakefulness (P < 0.05); ↑50%-90% ISF lactate (P < 0.05-0.001) | ↓Cognition | Enhances synaptic activity, promotes extracellular tau release, shortens tau half-life, and accelerates pathological deposition | Holth et al. (2019). Science[32] |
| Dietary Modulation | n-3 PUFAs (DHA) | fat-1 transgenic mice | N = 80-120 mice (total) + 6 human participants, n = 3-6 per group | ↑CSF outflow; ↑ISF solute clearance | ↓Aβ; ↓neuroinflammation; ↓neuronal damage | ↑CSF tracer (FITC-d3) influx and clearance in brain parenchyma (P < 0.05); ↓Aβ deposition in CA3 region (P < 0.05) | ↑Learning & memory | Improves AQP4 polarization | Ren et al. (2017). The FASEB Journal[130] |
| Dietary Modulation | High-doseFO | 5xFAD female mice (4 months) | N = 33-44 (total), n = 4-7 per group | ↓Aqp4 mRNA and protein expression | ↑Mfsd2a; ↓Aβ | ↑2.46-fold Mfsd2a mRNA (P < 0.001); ↑1.8-fold Mfsd2a vascular coverage (P < 0.001); ↓5.8-fold Aqp4 mRNA (P < 0.01); ↓3.2-fold Aqp4 protein (P < 0.001); ↓8.3-fold Lxrβ (P < 0.0001); ↓2.3-fold ABCA1 (P < 0.01); ↓21% Aβ accumulation in retinal vessels (P < 0.05); Srebp1-c normalized to control levels (P < 0.001) | / | FO-mediated upregulation of Mfsd2a and downregulation of AQP4, enhancing transcytosis and glymphatic drainage to reduce Aβ deposition. Mfsd2a restoration promotes DHA transport and inhibits caveolae formation, improving BRB integrity and Aβ clearance via transcytosis | Jovanovic et al. (2024). International Journal of Molecular Sciences[131] |
| Dietary Modulation | High-fat diet (HFD) | male C57BL/6J mice (3 weeks) | ↓CSF inflow;↓ISF solute clearance rate | ↑Aβ; ↑neuroinflammation; ↑neuronal damage | ↓Learning & memory | Long-term HFD disrupts the polarized distribution of AQP4 on astrocytic endfeet. This disruption directly impairs the function of the glymphatic system, leading to reduced clearance efficiency of Aβ and other waste products from the brain interstitium, thereby accelerating Aβ deposition and cognitive decline | Peng et al. (2025). Molecular Neurobiology[17] | ||
| Surgical Therapy | LVA | / | 6 AD patients | ↑CSF outflow | ↓Aβ; ↓Tau | Preliminary improvements in single case: ↑2 points MMSE (5→7); ↓2 points CDR-SB (10→8); ↓10.7% AD region T-sum (PET); ↑5.4% SUVmax; ↓global tau deposition; ↑90% ALPS index (1.19→2.26); | ↑Cognition & memory; ↓depression | / | Li et al. (2024). General Psychiatry[133] |
| Surgical Therapy | CBM | Adult male 5xFAD mice | N = 150-200 (total), n = 5-16 per group | ↑meningeal lymphatic vessel generation; ↑CSF drainage into DCLNs | ↓Aβ; ↓neuroinflammation | ↑30%-50% cognitive performance (NOR, Barnes maze, P < 0.05-0.001); ↓25%-50% Aβ deposition (P < 0.05-0.001); ↑30%-60% meningeal lymphangiogenesis (LYVE-1+ area, P < 0.05-0.001); ↑50%-80% lymphatic drainage to dCLNs (P < 0.01-0.001) | ↑Learning & memory | Through mechanical stress-induced cranial bone inflammation (IL-6 and VEGF-C upregulation) and meningeal lymphangiogenesis, which depend on the VEGF-C/VEGFR3 pathway to promote lymphatic endothelial cell proliferation and migration | Lu et al. (2025). Alzheimer’s & Dementia[137] |
CONCLUSION AND FUTURE PERSPECTIVES
The glymphatic system plays a pivotal role in AD by facilitating the clearance of neurotoxic proteins such as Aβ and tau. Its dysfunction, driven by aging, sleep disturbance, and genetic factors such as APOE4, accelerates neurodegeneration through impaired waste clearance, chronic neuroinflammation, and synaptic dysfunction.
Preclinical studies have identified multiple strategies to enhance glymphatic function, including pharmacological agents, TCM, neuromodulation, and lifestyle interventions. These approaches reduce pathological protein aggregation and improve cognitive outcomes in animal models. However, clinical translation remains challenging due to high failure rates in AD drug development, limitations of transgenic models, lack of standardized experimental protocols, and a scarcity of RCTs.
Despite these challenges, early-phase clinical studies have begun to explore the therapeutic potential of lymphatic-targeted interventions. A prospective single-arm trial evaluated deep cervical lymphatic-venous anastomosis in patients with severe AD, representing the first surgical approach directly targeting the lymphatic system in this context. The study reported preliminary evidence of cognitive stabilization and improvements in disease-related biomarkers (e.g., CSF Aβ42, p-tau181). In parallel, our group demonstrated that cerebellar iTBS, a non-invasive neuromodulation technique, enhances glymphatic clearance in AD patients. This effect was reflected by a significant increase in the DTI-ALPS index, which correlated with improved cognitive performance. Collectively, these findings offer initial clinical evidence supporting both surgical and non-invasive strategies for glymphatic enhancement[89,135,136].
Building on this emerging evidence base, future research should prioritize: (1) the development and validation of non-invasive imaging biomarkers for in vivo assessment of glymphatic function in humans; (2) well-designed, adequately powered RCTs to establish clinical efficacy; and (3) mechanistic studies investigating the interaction between glymphatic dysfunction and key neurodegenerative pathways. Advancing the field through rigorous, evidence-based methodologies will be essential to fully realize the therapeutic potential of glymphatic modulation in AD.
DECLARATIONS
Acknowledgments
The graphical abstract was created with BioRender.com [Created in BioRender. Niu, Y. (2026) https://BioRender.com/gi6maub].
Authors’ contributions
Conceptualized the study, developed the methodology, drafted the original manuscript, and supervised the project: Zhang X
Conducted the literature search, curated the data, and contributed to the original draft: Niu Y
Performed the investigation, formal analysis, and reviewed the manuscript: Wang X (Xiaomu Wang)
Validation, visualization, and manuscript review: Wang X (Xiaorui Wang)
Provided resources and participated in manuscript review: Li W
Handled software and reviewed the manuscript: Liu Z
Contributed to validation and manuscript review: Xu H
Managed project administration and reviewed the manuscript: Sun Z
Oversaw the study, acquired funding, and reviewed the manuscript: Jiang W
All authors read and approved the final manuscript.
Availability of data and materials
Not applicable.
AI and AI-assisted tool statement
Not applicable.
Financial support and sponsorship
This study was supported by the National Natural Science Foundation of China (grant number 82471471), Natural Science Basic Research Program of Shaanxi (grant number 2025JC-YBQN-1034), Shaanxi key research and development plan (grant number 2023-YBSF-353), the Joint Founding Project of Innovation Research Institute, Xijing Hospital (grant number LHJJ24JH13), Young Physician Cultivation Special Fund, Xijing Hospital, (grant number XJZT25QN46).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
2. Iliff JJ, Wang M, Liao Y, et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci Transl Med. 2012;4:147ra111.
3. Louveau A, Smirnov I, Keyes TJ, et al. Structural and functional features of central nervous system lymphatic vessels. Nature. 2015;523:337-41.
4. Huang SY, Zhang YR, Guo Y, et al. ; Alzheimer’s Disease Neuroimaging Initiative. Glymphatic system dysfunction predicts amyloid deposition, neurodegeneration, and clinical progression in Alzheimer’s disease. Alzheimers Dement. 2024;20:3251-69.
5. Nikolenko VN, Oganesyan MV, Vovkogon AD, et al. Current understanding of central nervous system drainage systems: implications in the context of neurodegenerative diseases. Curr Neuropharmacol. 2020;18:1054-63.
6. Shan X, Lu Y, Luo Z, et al. A long-acting lyotropic liquid crystalline implant promotes the drainage of macromolecules by brain-related lymphatic system in treating aged Alzheimer’s disease. ACS Nano. 2024;18:9688-703.
7. Li Y, Zhang J, Wan J, Liu A, Sun J. Melatonin regulates Aβ production/clearance balance and Aβ neurotoxicity: a potential therapeutic molecule for Alzheimer’s disease. Biomed Pharmacother. 2020;132:110887.
9. Song YY, Xu WT, Zhang XC, Ni GX. Mechanisms of electroacupuncture on Alzheimer’s Disease: a review of animal studies. Chin J Integr Med. 2020;26:473-80.
10. Lin Y, Jin J, Lv R, et al. Repetitive transcranial magnetic stimulation increases the brain’s drainage efficiency in a mouse model of Alzheimer’s disease. Acta Neuropathol Commun. 2021;9:102.
11. Ju YS, Ooms SJ, Sutphen C, et al. Slow wave sleep disruption increases cerebrospinal fluid amyloid-β levels. Brain. 2017;140:2104-11.
12. Hablitz LM, Nedergaard M. The glymphatic system: a novel component of fundamental neurobiology. J Neurosci. 2021;41:7698-711.
13. Formolo DA, Yu J, Lin K, et al. Leveraging the glymphatic and meningeal lymphatic systems as therapeutic strategies in Alzheimer’s disease: an updated overview of nonpharmacological therapies. Mol Neurodegener. 2023;18:26.
14. Lohela TJ, Lilius TO, Nedergaard M. The glymphatic system: implications for drugs for central nervous system diseases. Nat Rev Drug Discov. 2022;21:763-79.
15. Li Y, Meng Q, Luo B, et al. Exercises in activating lymphatic system on fluid overload symptoms, abnormal weight gains, and physical functions among patients with heart failure: a randomized controlled trial. Front Cardiovasc Med. 2023;10:1094805.
16. Togre NS, Mekala N, Bhoj PS, et al. Neuroinflammatory responses and blood-brain barrier injury in chronic alcohol exposure: role of purinergic P2 × 7 Receptor signaling. J Neuroinflammation. 2024;21:244.
17. Peng W, Yuan Y, Lei J, et al. Long-term high-fat diet impairs AQP4-mediated glymphatic clearance of amyloid beta. Mol Neurobiol. 2025;62:1079-93.
18. Xie L, Kang H, Xu Q, et al. Sleep drives metabolite clearance from the adult brain. Science. 2013;342:373-7.
19. Jiang-Xie LF, Drieu A, Kipnis J. Waste clearance shapes aging brain health. Neuron. 2025;113:71-81.
20. Da Mesquita S, Louveau A, Vaccari A, et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature. 2018;560:185-91.
21. Da Mesquita S, Papadopoulos Z, Dykstra T, et al. ; Dominantly Inherited Alzheimer Network. Meningeal lymphatics affect microglia responses and anti-Aβ immunotherapy. Nature. 2021;593:255-60.
22. Hong H, Hong L, Luo X, et al. ; Alzheimer’s Disease Neuroimaging Initiative (ADNI). The relationship between amyloid pathology, cerebral small vessel disease, glymphatic dysfunction, and cognition: a study based on Alzheimer’s disease continuum participants. Alzheimers Res Ther. 2024;16:43.
23. Chachaj A, Gąsiorowski K, Szuba A, Sieradzki A, Leszek J. The lymphatic system in the brain clearance mechanisms - new therapeutic perspectives for Alzheimer’s disease. Curr Neuropharmacol. 2023;21:380-91.
24. Prince M, Bryce R, Albanese E, Wimo A, Ribeiro W, Ferri CP. The global prevalence of dementia: a systematic review and metaanalysis. Alzheimers Dement. 2013;9:63.
25. Kress BT, Iliff JJ, Xia M, et al. Impairment of paravascular clearance pathways in the aging brain. Ann Neurol. 2014;76:845-61.
26. Beschorner N, Nedergaard M. Glymphatic system dysfunction in neurodegenerative diseases. Curr Opin Neurol. 2024;37:182-8.
27. Zhang S, Gao Y, Zhao Y, Huang TY, Zheng Q, Wang X. Peripheral and central neuroimmune mechanisms in Alzheimer’s disease pathogenesis. Mol Neurodegener. 2025;20:22.
28. Plog BA, Dashnaw ML, Hitomi E, et al. Biomarkers of traumatic injury are transported from brain to blood via the glymphatic system. J Neurosci. 2015;35:518-26.
29. Lundgaard I, Lu ML, Yang E, et al. Glymphatic clearance controls state-dependent changes in brain lactate concentration. J Cereb Blood Flow Metab. 2017;37:2112-24.
30. Guo C, Harshfield EL, Markus HS. Sleep characteristics and risk of stroke and dementia: an observational and mendelian randomization study. Neurology. 2024;102:e209141.
31. Yaffe K, Falvey CM, Hoang T. Connections between sleep and cognition in older adults. Lancet Neurol. 2014;13:1017-28.
32. Holth JK, Fritschi SK, Wang C, et al. The sleep-wake cycle regulates brain interstitial fluid tau in mice and CSF tau in humans. Science. 2019;363:880-4.
33. Kang J, Lim MM, Bateman RJ, et al. Amyloid-beta dynamics are regulated by orexin and the sleep-wake cycle. Science. 2009;326:1005-7.
34. Kiviniemi V, Wang X, Korhonen V, et al. Ultra-fast magnetic resonance encephalography of physiological brain activity - Glymphatic pulsation mechanisms? J Cereb Blood Flow Metab. 2016;36:1033-45.
35. Ringstad G, Valnes LM, Dale AM, et al. Brain-wide glymphatic enhancement and clearance in humans assessed with MRI. JCI Insight. 2018;3:e121537.
36. Zhu Q, Song Y, Qian Y, Zhang R, Xue J, Hou Y. Targeting glial dysfunction in Alzheimer’s disease: insights into pathogenesis and emerging therapeutics. Ageing Neur Dis. 2025;5:19.
37. Chen X, Firulyova M, Manis M, et al. Microglia-mediated T cell infiltration drives neurodegeneration in tauopathy. Nature. 2023;615:668-77.
38. Kaya T, Mattugini N, Liu L, et al. CD8+ T cells induce interferon-responsive oligodendrocytes and microglia in white matter aging. Nat Neurosci. 2022;25:1446-57.
39. Plantone D, Pardini M, Locci S, Nobili F, De Stefano N. B lymphocytes in Alzheimer’s disease - a comprehensive review. J of Alzheimers Dis. 2022;88:1241-62.
40. Paouri E, Georgopoulos S. Systemic and CNS inflammation crosstalk: implications for Alzheimer’s disease. Curr Alzheimer Res. 2019;16:559-74.
41. Wu J, Gao G, Shi F, et al. Activated microglia-induced neuroinflammatory cytokines lead to photoreceptor apoptosis in Aβ-injected mice. J Mol Med. 2021;99:713-28.
42. Unger M, Li E, Scharnagl L, et al. CD8+ T-cells infiltrate Alzheimer’s disease brains and regulate neuronal- and synapse-related gene expression in APP-PS1 transgenic mice. Brain Behav Immun. 2020;89:67-86.
44. Bettcher BM, Tansey MG, Dorothée G, Heneka MT. Peripheral and central immune system crosstalk in Alzheimer disease - a research prospectus. Nat Rev Neurol. 2021;17:689-701.
45. Wang L, Zhang Y, Zhao Y, Marshall C, Wu T, Xiao M. Deep cervical lymph node ligation aggravates AD‐like pathology of APP/PS1 mice. Brain Pathol. 2018;29:176-92.
46. Kim K, Abramishvili D, Du S, et al. Meningeal lymphatics-microglia axis regulates synaptic physiology. Cell. 2025;188:2705-19.e23.
47. Tavares GA, Louveau A. Meningeal lymphatics: an immune gateway for the central nervous system. Cells. 2021;10:3385.
48. Smith AJ, Duan T, Verkman AS. Aquaporin-4 reduces neuropathology in a mouse model of Alzheimer’s disease by remodeling peri-plaque astrocyte structure. Acta Neuropathol Commun. 2019;7:74.
49. Mestre H, Hablitz LM, Xavier AL, et al. Aquaporin-4-dependent glymphatic solute transport in the rodent brain. ELife. 2018;7:e40070.
50. Yang J, Lunde LK, Nuntagij P, et al. Loss of astrocyte polarization in the tg-ArcSwe mouse model of Alzheimer’s disease. J Alzheimers Dis. 2011;27:711-22.
51. Zeppenfeld DM, Simon M, Haswell JD, et al. Association of perivascular localization of aquaporin-4 with cognition and Alzheimer disease in aging brains. JAMA Neurol. 2017;74:91-9.
52. Mentis AA, Dardiotis E, Chrousos GP. Apolipoprotein E4 and meningeal lymphatics in Alzheimer disease: a conceptual framework. Mol Psychiatry. 2020;26:1075-97.
53. Raulin A, Doss SV, Trottier ZA, Ikezu TC, Bu G, Liu C. ApoE in Alzheimer’s disease: pathophysiology and therapeutic strategies. Mol Neurodegener. 2022;17:72.
54. Kaji S, Berghoff SA, Spieth L, et al. Apolipoprotein E aggregation in microglia initiates Alzheimer’s disease pathology by seeding β-amyloidosis. Immunity. 2024;57:2651-68.e12.
55. Bell RD, Winkler EA, Singh I, et al. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature. 2012;485:512-6.
56. Kang KM, Park C, Byun MS, et al. ; KBASE Research Group. APOE4 modulates the association between DTI‐ALPS index and Alzheimer’s pathologies. Alzheimers Dement. 2025;21:e70837.
57. Achariyar TM, Li B, Peng W, et al. Glymphatic distribution of CSF-derived apoE into brain is isoform specific and suppressed during sleep deprivation. Mol Neurodegener. 2016;11:74.
58. Devenish SRA. The current landscape in Alzheimer’s disease research and drug discovery. Drug Discov Today. 2020;25:943-5.
59. Lyu D, Lyu X, Huang L, Fang B. Effects of three kinds of anti-amyloid-β drugs on clinical, biomarker, neuroimaging outcomes and safety indexes: a systematic review and meta-analysis of phase II/III clinical trials in Alzheimer’s disease. Ageing Res Rev. 2023;88:101959.
60. Yeo-Teh NSL, Tang BL. A review of scientific ethics issues associated with the recently approved drugs for Alzheimer’s disease. Sci Eng Ethics. 2023;29:2.
61. Wu YH, Feenstra MGP, Zhou JN, et al. Molecular changes underlying reduced pineal melatonin levels in Alzheimer disease: alterations in preclinical and clinical stages. J Clin Endocrinol Metab. 2003;88:5898-906.
62. Pappolla MA, Matsubara E, Vidal R, et al. Melatonin treatment enhances Aβ lymphatic clearance in a transgenic mouse model of amyloidosis. Curr Alzheimer Res. 2018;15:637-42.
63. Wade AG, Farmer M, Harari G, et al. Add-on prolonged-release melatonin for cognitive function and sleep in mild to moderate Alzheimer’s disease: a 6-month, randomized, placebo-controlled, multicenter trial. Clin Interv Aging. 2014;9:94761.
64. Manescu MD, Catalin B, Baldea I, et al. Aquaporin 4 modulation drives amyloid burden and cognitive abilities in an APPPS1 mouse model of Alzheimer’s disease. Alzheimers Dement. 2025;21:e70164.
65. Sasaki K, Fujita H, Sato T, et al. GLP-1 receptor signaling restores aquaporin 4 subcellular polarization in reactive astrocytes and promotes amyloid β clearance in a mouse model of Alzheimer’s disease. Biochem Biophys Res Commun. 2024;741:151016.
66. Zhang B, Li W, Zhuo Y, et al. L-3-n-butylphthalide effectively improves the glymphatic clearance and reduce amyloid-β deposition in Alzheimer’s transgenic mice. J Mol Neurosci. 2021;71:1266-74.
67. Zhang X, Cao R, Zhu C, et al. Mechanism of anti-AD action of OAB-14 by enhancing the function of glymphatic system. Neurochem Int. 2023;171:105633.
68. Ye C, Wang S, Niu L, et al. Unlocking potential of oxytocin: improving intracranial lymphatic drainage for Alzheimer’s disease treatment. Theranostics. 2024;14:4331-51.
69. Maki T, Okamoto Y, Carare RO, et al. Phosphodiesterase III inhibitor promotes drainage of cerebrovascular β‐amyloid. Ann Clin Transl Neurol. 2014;1:519-33.
70. Kimura T, Hamazaki TS, Sugaya M, et al. Cilostazol improves lymphatic function by inducing proliferation and stabilization of lymphatic endothelial cells. J Dermatol Sci. 2014;74:150-8.
71. Aspelund A, Antila S, Proulx ST, et al. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J Exp Med. 2015;212:991-9.
72. Antila S, Karaman S, Nurmi H, et al. Development and plasticity of meningeal lymphatic vessels. J Exp Med. 2017;214:3645-67.
73. Plog BA, Mestre H, Olveda GE, et al. Transcranial optical imaging reveals a pathway for optimizing the delivery of immunotherapeutics to the brain. JCI Insight. 2018;3:e126138.
74. Lilius TO, Blomqvist K, Hauglund NL, et al. Dexmedetomidine enhances glymphatic brain delivery of intrathecally administered drugs. J Controlled Release. 2019;304:29-38.
75. Lopes DM, Wells JA, Ma D, et al. Glymphatic inhibition exacerbates tau propagation in an Alzheimer’s disease model. Alzheimers Res Ther. 2024;16:71.
76. Wang J, Wong YK, Liao F. What has traditional Chinese medicine delivered for modern medicine? Expert Rev Mol Med. 2018;20:e4.
77. Wu Y, Zhang T, Li X, et al. Borneol-driven meningeal lymphatic drainage clears amyloid-β peptide to attenuate Alzheimer-like phenotype in mice. Theranostics. 2023;13:106-24.
78. Li J, Hao Y, Wang S, et al. Yuanzhi powder facilitated Aβ clearance in APP/PS1 mice: Target to the drainage of glymphatic system and meningeal lymphatic vessels. J Ethnopharmacol. 2024;319:117195.
79. Liu X, Zhang H, Xiang J, et al. Jiawei Xionggui Decoction promotes meningeal lymphatic vessels clearance of β-amyloid by inhibiting arachidonic acid pathway. Phytomedicine. 2024;135:156041.
80. Wang FJ, Wang SX, Chai LJ, Zhang Y, Guo H, Hu LM. Xueshuantong injection (lyophilized) combined with salvianolate lyophilized injection protects against focal cerebral ischemia/reperfusion injury in rats through attenuation of oxidative stress. Acta Pharmacol Sin. 2017;39:998-1011.
81. Zheng R, Huang YM, Zhou Q. Xueshuantong improves functions of lymphatic ducts and modulates inflammatory responses in Alzheimer’s disease mice. Front Pharmacol. 2021;12:605814.
82. Song B, Ao Q, Niu Y, et al. Amyloid beta-peptide worsens cognitive impairment following cerebral ischemia-reperfusion injury. Neural Regen Res 2013;8:2449-57. DOI: 10.3969/j.issn.1673-5374.2013.26.006.
83. Wang B, Lyu Z, Chan Y, et al. Tongxinluo exerts inhibitory effects on pyroptosis and amyloid-β peptide accumulation after cerebral ischemia/reperfusion in rats. Evid Based Complement Alternat Med. 2021;2021:5788602.
84. Lyu Z, Li Q, Yu Z, et al. Yi-Zhi-Fang-Dai formula exerts neuroprotective effects against pyroptosis and blood-brain barrier-glymphatic dysfunctions to prevent amyloid-beta acute accumulation after cerebral ischemia and reperfusion in rats. Front Pharmacol. 2021;12:791059.
85. Okamoto M, Mizuuchi D, Omura K, et al. High-intensity intermittent training enhances spatial memory and hippocampal neurogenesis associated with BDNF signaling in rats. Cereb Cortex. 2021;31:4386-97.
86. Van Reet J, Tunnell K, Anderson K, et al. Evaluation of advective solute infiltration into porous media by pulsed focused ultrasound-induced acoustic streaming effects. Ultrasonography. 2024;43:35-46.
87. Lee Y, Choi Y, Park EJ, et al. Improvement of glymphatic-lymphatic drainage of beta-amyloid by focused ultrasound in Alzheimer’s disease model. Sci Rep. 2020;10:16144.
88. Mehta RI, Carpenter JS, Mehta RI, et al. Ultrasound-mediated blood-brain barrier opening uncovers an intracerebral perivenous fluid network in persons with Alzheimer’s disease. Fluids Barriers CNS. 2023;20:46.
89. Zhang X, Sun Z, Wu D, et al. Effects of cerebellar intermittent theta-burst stimulation on patients with Alzheimer’s disease: A randomized controlled trial. Journal of Alzheimer’s Disease. 2025;107:1187-99.
90. Xia KP, Pang J, Li SL, Zhang M, Li HL, Wang YJ. Effect of electroacupuncture at governor vessel on learning-memory ability and serum level of APP, Aβ1-42 in patients with Alzheimer’s disease. Zhongguo Zhen Jiu. 2020;40:375-8.
91. Feng Q, Bin LL, Zhai YB, Xu M, Liu ZS, Peng WN. Long-term efficacy and safety of electroacupuncture on improving MMSE in patients with Alzheimer’s disease. Zhongguo Zhen Jiu. 2019;39:3-8.
92. Liang P, Li L, Zhang Y, et al. Electroacupuncture Improves Clearance of Amyloid-β through the Glymphatic System in the SAMP8 Mouse Model of Alzheimer’s Disease. Neural Plast. 2021;2021:1-11.
93. Wang LM, Zhao TT, Zhou HP, et al. Effect of electroacupuncture on recognition memory and levels of Aβ, inflammatory factor proteins and aquaporin 4 in hippocampus of APP/PS1 double transgenic mice. Zhen Ci Yan Jiu. 2020;45:431-7.
94. Glass GE. Photobiomodulation: a review of the molecular evidence for low level light therapy. J Plast Reconstr Aesthet Surg. 2021;74:1050-60.
95. Santana-Blank L, Rodríguez-Santana E, Santana-Rodríguez K. Theoretic, experimental, clinical bases of the water oscillator hypothesis in near-infrared photobiomodulation. Photomed Laser Surg. 2010;28:S41-52.
96. de Freitas LF, Hamblin MR. Proposed mechanisms of photobiomodulation or low-level light therapy. IEEE J Sel Top Quantum Electron. 2016;22:37000417.
97. Ganeshan V, Skladnev NV, Kim JY, Mitrofanis J, Stone J, Johnstone DM. Pre-conditioning with remote photobiomodulation modulates the brain transcriptome and protects against MPTP insult in mice. Neuroscience. 2019;400:85-97.
98. George S, Hamblin MR, Abrahamse H. Photobiomodulation‐induced differentiation of immortalized adipose stem cells to neuronal cells. Lasers Surg Med. 2020;52:1032-40.
99. Shen Q, Guo H, Yan Y. Photobiomodulation for neurodegenerative diseases: a scoping review. Int J Mol Sci. 2024;25:1625.
100. Zinchenko E, Navolokin N, Shirokov A, et al. Pilot study of transcranial photobiomodulation of lymphatic clearance of beta-amyloid from the mouse brain: breakthrough strategies for non-pharmacologic therapy of Alzheimer’s disease. Biomed Opt Express. 2019;10:4003-17.
101. Semyachkina-Glushkovskaya O, Klimova M, Iskra T, et al. Transcranial photobiomodulation of clearance of beta-amyloid from the mouse brain: effects on the meningeal lymphatic drainage and blood oxygen saturation of the brain. Adv Exp Med Biol. 2021;1269:57-61.
102. Wang M, Yan C, Li X, et al. Non-invasive modulation of meningeal lymphatics ameliorates ageing and Alzheimer’s disease-associated pathology and cognition in mice. Nat Commun. 2024;15:1453.
103. Wu W, Zhao Y, Cheng X, et al. Modulation of glymphatic system by visual circuit activation alleviates memory impairment and apathy in a mouse model of Alzheimer’s disease. Nat Commun. 2025;16:63.
104. Lian X, Liu Z, Gan Z, et al. Targeting the glymphatic system to promote α-synuclein clearance: a novel therapeutic strategy for Parkinson’s disease. Neural Regen Res. 2026;21:233-47.
105. Vedam-Mai V, Van Battum EY, Kamphuis W, et al. Deep brain stimulation and the role of astrocytes. Mol Psychiatry. 2012;17:124-31.
106. Jang JS, Choi C, Yi J, et al. High frequency electrical stimulation promotes expression of extracellular matrix proteins from human astrocytes. Mol Biol Rep. 2019;46:4369-75.
107. Naour AL, Beziat E, Kam JH, Magistretti P, Benabid AL, Mitrofanis J. Do astrocytes respond to light, sound, or electrical stimulation? Neural Regen Res. 2023;18:2343-7.
108. Wang Y, Zhan G, Cai Z, et al. Vagus nerve stimulation in brain diseases: therapeutic applications and biological mechanisms. Neurosci Biobehav Rev. 2021;127:37-53.
109. Cheng KP, Brodnick SK, Blanz SL, et al. Clinically-derived vagus nerve stimulation enhances cerebrospinal fluid penetrance. Brain Stimul. 2020;13:1024-30.
110. Chase HW, Boudewyn MA, Carter CS, Phillips ML. Transcranial direct current stimulation: a roadmap for research, from mechanism of action to clinical implementation. Mol Psychiatry. 2019;25:397-407.
111. Wang Y, Monai H. Transcranial direct current stimulation alters cerebrospinal fluid-interstitial fluid exchange in mouse brain. Brain Stimul. 2024;17:620-32.
112. Huuha AM, Norevik CS, Moreira JBN, et al. Can exercise training teach us how to treat Alzheimer’s disease? Ageing Res Rev. 2022;75:101559.
113. Li S. Benefits of physical exercise on Alzheimer’s disease: an epigenetic view. Ageing Neur Dis. 2023;3:6.
114. Lee TH, Formolo DA, Kong T, et al. Potential exerkines for physical exercise-elicited pro-cognitive effects: Insight from clinical and animal research. Int Rev Neurobiol. 2019;147:361-95.
115. Guure CB, Ibrahim NA, Adam MB, Said SM. Impact of physical activity on cognitive decline, dementia, and its subtypes: meta-analysis of prospective studies. Biomed Res Int. 2017;2017:9016924.
116. Khodadadi D, Gharakhanlou R, Naghdi N, et al. Treadmill exercise ameliorates spatial learning and memory deficits through improving the clearance of peripheral and central amyloid-beta levels. Neurochem Res. 2018;43:1561-74.
117. He XF, Liu DX, Zhang Q, et al. Voluntary exercise promotes glymphatic clearance of amyloid beta and reduces the activation of astrocytes and microglia in aged mice. Front Mol Neurosci. 2017;10:144.
118. von Holstein-Rathlou S, Petersen NC, Nedergaard M. Voluntary running enhances glymphatic influx in awake behaving, young mice. Neurosci Lett. 2018;662:253-8.
119. Liang S, Liu H, Wang X, et al. Aerobic exercise improves clearance of amyloid-β via the glymphatic system in a mouse model of Alzheimer’s disease. Brain Res Bull. 2025;222:111263.
120. Liu Y, Hu PP, Zhai S, et al. Aquaporin 4 deficiency eliminates the beneficial effects of voluntary exercise in a mouse model of Alzheimer’s disease. Neural Regen Res. 2022;17:2079-88.
121. Li M, Xu J, Li L, et al. Voluntary wheel exercise improves glymphatic clearance and ameliorates colitis-associated cognitive impairment in aged mice by inhibiting TRPV4-induced astrocytic calcium activity. Exp Neurol. 2024;376:114770.
122. Gholipour P, Komaki A, Ramezani M, Parsa H. Effects of the combination of high-intensity interval training and ecdysterone on learning and memory abilities, antioxidant enzyme activities, and neuronal population in an amyloid-beta-induced rat model of Alzheimer’s disease. Physiol Behav. 2022;251:113817.
123. Feng S, Wu C, Zou P, et al. High-intensity interval training ameliorates Alzheimer’s disease-like pathology by regulating astrocyte phenotype-associated AQP4 polarization. Theranostics. 2023;13:3434-50.
124. Ma Y, Liang L, Zheng F, Shi L, Zhong B, Xie W. Association between sleep duration and cognitive decline. JAMA Netw Open. 2020;3:e2013573.
125. Benca R, Herring WJ, Khandker R, Qureshi ZP. Burden of insomnia and sleep disturbances and the impact of sleep treatments in patients with probable or possible Alzheimer’s disease: a structured literature review. J Alzheimers Dis. 2022;86:83-109.
126. Turton SM, Padgett S, Maisel MT, et al. Interactions between daily sleep-wake rhythms, γ-secretase, and amyloid-β peptide pathology point to complex underlying relationships. Biochim Biophys Acta Mol Basis Dis. 2025;1871:167840.
127. Thipani Madhu M, Balaji O, Kandi V, et al. Role of the glymphatic system in Alzheimer’s disease and treatment approaches: a narrative review. Cureus. 2024;16:e63448.
128. Pistollato F, Iglesias RC, Ruiz R, et al. Nutritional patterns associated with the maintenance of neurocognitive functions and the risk of dementia and Alzheimer’s disease: a focus on human studies. Pharmacol Res. 2018;131:32-43.
129. Wen J, Satyanarayanan SK, Li A, et al. Unraveling the impact of Omega-3 polyunsaturated fatty acids on blood-brain barrier (BBB) integrity and glymphatic function. Brain Behav Immun. 2024;115:335-55.
130. Ren H, Luo C, Feng Y, et al. Omega‐3 polyunsaturated fatty acids promote amyloid‐β clearance from the brain through mediating the function of the glymphatic system. FASEB J. 2017;31:282-93.
131. Jovanovic Macura I, Milanovic D, Tesic V, et al. The impact of high-dose fish oil supplementation on Mfsd2a, Aqp4, and amyloid-β expression in retinal blood vessels of 5xFAD Alzheimer’s mouse model. Int J Mol Sci. 2024;25:9400.
132. Xie Q, Louveau A, Pandey S, Zeng W, Chen WF. Rewiring the brain: the next frontier in supermicrosurgery. Plast Reconstr Surg. 2023;153:494e-5e.
133. Li X, Zhang C, Fang Y, et al. Promising outcomes 5 weeks after a surgical cervical shunting procedure to unclog cerebral lymphatic systems in a patient with Alzheimer’s disease. Gen Psychiatr. 2024;37:e101641.
134. Fang R, Jin L, Lu H, Xie Q, Yang X, Kueckelhaus M. A novel microsurgical model of cervical lymph node-to-vein anastomosis (LNVA) for studying brain lymphatic outflow. J Craniofac Surg. 2025;36:2160-3.
135. Chen JY, Zhao DW, Yin Y, et al. Deep cervical lymphovenous anastomosis (LVA) for Alzheimer’s disease: microsurgical procedure in a prospective cohort study. Int J Surg. 2025;111:4211-21.
136. Fu X, Zhang J, Xiao Q, et al. Deep cervical lymphatic-venous anastomosis attenuates cognitive dysfunction and biomarker abnormalities in severe Alzheimer’s disease: a prospective single‐arm study. Alzheimers Dement. 2026;22:e71150.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0













Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].