


The PFAS-lipid-frailty nexus: an interconnected pathway to healthy aging
 0
0 
INTRODUCTION
Global population aging presents immense challenges and opportunities for sustainable development. Frailty, a state of increased vulnerability to adverse health outcomes due to age-related decline in multiple physiological systems, affects a substantial proportion of older adults and is a major predictor of disability, hospitalization, and mortality[1,2]. Its pathogenesis involves chronic low-grade inflammation (“inflammaging”), oxidative stress, mitochondrial dysfunction, and metabolic dysregulation[3-6]. While nutritional factors like omega-3 fatty acids are recognized as protective[7,8], the contribution of ubiquitous environmental pollutants to frailty remains underexplored.
Per- and polyfluoroalkyl substances (PFAS) are a large class of persistent synthetic chemicals that have been used for decades in consumer and industrial products. Their extreme environmental persistence and widespread detection in human populations have raised concerns about health effects[9,10]. PFAS exposure is associated with dyslipidemia, hepatotoxicity, immune suppression, and endocrine disruption[11,12]. Mechanistically, PFAS can induce oxidative stress and mitochondrial damage, and activate nuclear receptors, particularly peroxisome proliferator-activated receptor alpha (PPARα), a master regulator of lipid metabolism[13,14]. These pathways strikingly overlap with the core biological processes underlying frailty, suggesting a plausible, yet largely uninvestigated, link.
In this opinion, we propose a conceptual framework connecting PFAS exposure to frailty risk, with the omega-3 fatty acids docosahexaenoic acid (DHA)/eicosapentaenoic acid (EPA) and the lysophosphatidylcholine (LPC) lipid axis as key intermediaries. By integrating evidence across environmental health, nutrition, and lipid biology, we aim to stimulate transdisciplinary research and inform potential prevention strategies within a “One Health” framework.
PFAS, LIPID METABOLISM, AND THE NEXUS WITH FRAILTY BIOLOGY
PFAS are recognized for their potent ability to perturb lipid homeostasis. Epidemiological studies consistently associate PFAS exposure with altered serum lipid profiles, including increased total and non-high-density lipoprotein (HDL) cholesterol[15]. Metabolomics studies have further revealed that PFAS exposure is associated with widespread disruption of lipid-related metabolites, including glycerophospholipids and fatty acids[16,17], providing a systems-level view of this metabolic interference. The liver is a primary target organ for PFAS accumulation and toxicity. PFAS activate PPARα, leading to alterations in genes involved in fatty acid uptake, β-oxidation, and lipoprotein synthesis[13,18]. However, PPARα activation alone cannot explain the full spectrum of PFAS-induced lipid changes; other nuclear receptors [constitutive androstane receptor (CAR), pregnane X receptor (PXR)], oxidative stress, and direct mitochondrial interference also contribute[14,19]. Moreover, interspecies differences in PPARα sensitivity - rodents are generally more responsive than humans - caution against direct extrapolation of animal data[20,21]. Nevertheless, humanized PPARα mouse models show that even when PPARα responses are dampened, PFOA still affects cholesterol homeostasis, suggesting additional mechanisms are at play[21]. Additionally, concurrent exposure to other metabolism-disrupting chemicals [e.g., bisphenol A (BPA), phthalates, persistent organic pollutants (POPs)] may produce additive or synergistic effects through shared pathways[22,23]. These complexities must be considered when linking PFAS to frailty.
Importantly, emerging evidence suggests that even some short-chain replacement PFAS can also significantly impact hepatic lipid droplet accumulation[24,25], underscoring the need to assess a broad range of PFAS, including novel substitutes such as 6:2 fluorotelomer sulfonic acid (6:2 FTSA), which exhibit distinct environmental transformation profiles[26].
These PFAS-induced perturbations in lipid metabolism - particularly in the liver - could have systemic consequences that resonate with frailty mechanisms. Dysregulated fatty acid metabolism can fuel oxidative stress and inflammation[27]. Altered hepatic production of lipid mediators and lipoproteins could influence peripheral tissues, including skeletal muscle, where lipid accumulation and mitochondrial dysfunction are hallmarks of sarcopenia and frailty[4,28]. However, skeletal muscle is not merely a passive recipient of liver-derived signals; PFAS can accumulate directly in muscle, perturb oxidative phosphorylation, reduce lean mass, and induce local mitochondrial dysfunction[29-31]. Thus, the link from hepatic remodeling to muscle decline likely involves both systemic LPC-mediated effects and direct muscle toxicity.
DHA/EPA: PROTECTIVE NUTRIENTS AT THE INTERFACE OF ENVIRONMENT AND AGING
DHA and EPA are long-chain omega-3 polyunsaturated fatty acids (PUFAs) with well-documented anti-inflammatory and pro-resolving properties[32,33]. They are incorporated into cell membrane phospholipids, where they influence membrane fluidity, lipid raft organization, and serve as precursors for specialized pro-resolving mediators (SPMs) such as resolvins and protectins[34,35].
Epidemiological evidence links higher omega-3 status with better physical function and lower frailty risk. A recent cohort study reported that higher erythrocyte levels of n-3 PUFAs, particularly DHA, were associated with reduced incidence of frailty in older adults[36]. Randomized controlled trials, such as the large European DO-HEALTH trial (Vitamin D3–Omega-3–Home Exercise–Healthy Aging and Longevity Trial; 2 × 2 × 2 factorial design, 2,157 participants ≥ 70 years), provide preliminary evidence that omega-3 supplementation (1 g/day), alone or combined with vitamin D and exercise, may have beneficial effects on pre-frailty and other aging-related outcomes[7,37]. The mechanisms are thought to involve reduced inflammation, improved muscle protein synthesis, and enhanced mitochondrial function[7,8]. Importantly, effects likely depend on baseline nutritional status, dose, duration, and co-interventions[7].
Crucially, the incorporation of DHA/EPA into membranes and their subsequent functions are intricately linked to the pathways that PFAS disrupt. PFAS activation of PPARα can influence the expression of enzymes involved in PUFA metabolism and phospholipid incorporation[38]. Thus, PFAS exposure could interfere with the availability or membrane integration of these protective fatty acids, diminishing their beneficial effects. It should also be noted that seafood is a dual source of both omega-3s and PFAS, which may confound epidemiological associations[39].
THE LPC LIPID AXIS: A MECHANISTIC HUB CONNECTING PFAS, FATTY ACIDS, AND FRAILTY
LPCs are a class of bioactive lipids generated primarily from phosphatidylcholine (PC) by phospholipase A2 (PLA2) enzymes. They can be re-esterified back to PC by lysophosphatidylcholine acyltransferases (LPCATs) in the continuous Lands’ cycle, which is fundamental for membrane phospholipid remodeling[40]. This cycle dynamically controls the asymmetric distribution and turnover of fatty acids, including PUFAs such as DHA and arachidonic acid, within membrane PC pools[40]. The LPCAT enzymes, particularly LPCAT3, play a critical role in enriching membranes with PUFAs, thereby influencing membrane properties and the availability of substrate for eicosanoid and SPM synthesis[39]. This pathway is especially relevant to aging muscle, which relies on efficient membrane remodeling for contractile function and mitochondrial integrity.
Emerging evidence positions LPCs as potential biomarkers and mediators in frailty. Metabolomic studies have repeatedly identified specific LPC species (e.g., LPC-DHA, LPC-EPA) that are lower in frail individuals compared to robust older adults, and these alterations correlate with muscle strength and mass[41,42]. A recent study in a frailty clinic setting found that lower plasma LPC levels were associated with reduced handgrip strength[43]. While these cross-sectional associations are intriguing, they do not establish causality; altered LPC profiles may also reflect malnutrition, inflammation, liver function, or existing frailty[44]. Nevertheless, the consistent signal suggests that the LPC lipid axis is functionally linked to muscle health.
We propose that the LPC lipid axis serves as a critical mechanistic bridge connecting PFAS exposure, DHA/EPA status, and frailty. In this framework [Figure 1]:
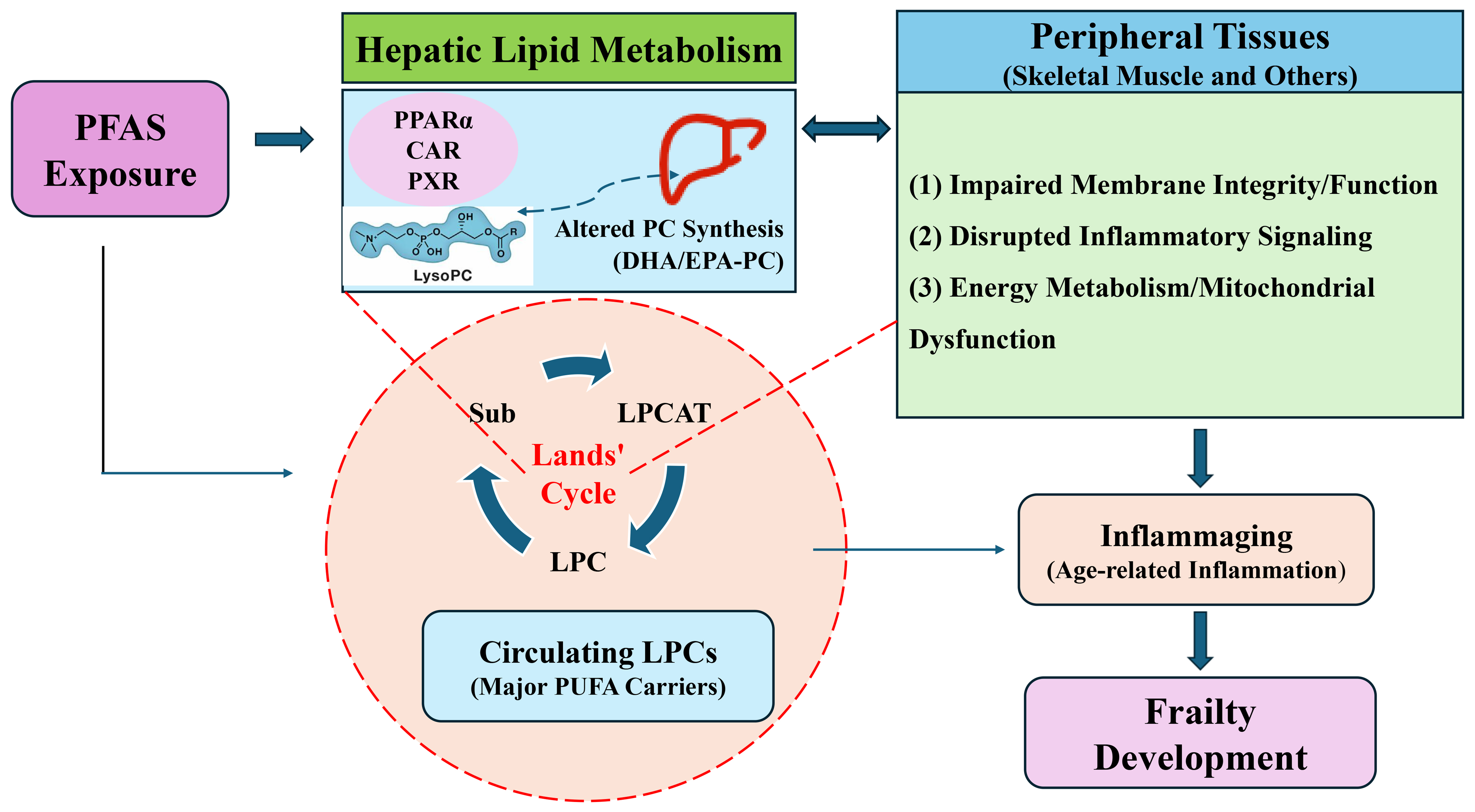
Figure 1. Conceptual framework illustrating the hypothesized PFAS-lipid-frailty nexus. The LIVER icon was adapted from Iconfont (https://www.iconfont.cn/). PFAS are proposed to disrupt hepatic lipid metabolism via PPARα and other pathways, altering the pool of DHA/EPA-containing PCs. This changes the substrate for PLA2, impacting the production of specific LPCs within the Lands’ cycle (LPCAT-mediated reacylation). The resulting alteration in the circulating LPC profile affects the delivery of DHA/EPA to peripheral tissues (e.g., skeletal muscle), influencing membrane function, inflammatory tone, and energy metabolism. These disruptions, superimposed on age-related inflammaging, contribute to frailty pathophysiology. Direct PFAS toxicity in muscle and other tissues may also occur. The pathways depicted are hypothetical and require empirical validation. PFAS: Per- and polyfluoroalkyl substances; PPARα: peroxisome proliferator-activated receptor alpha; DHA: docosahexaenoic acid; EPA: eicosapentaenoic acid; PCs: phosphatidylcholines; PLA2: phospholipase A2; LPCs: lysophosphatidylcholines; LPCAT: lysophosphatidylcholine acyltransferase; CAR: constitutive androstane receptor; PXR: pregnane X receptor; Sub: Substrate; PUFA: polyunsaturated fatty acid.
PFAS exposure disrupts hepatic lipid metabolism via PPARα and other pathways, altering the synthesis and composition of PCs, including those containing DHA and EPA. This altered substrate pool changes the availability of DHA/EPA-PCs as substrates for PLA2, thereby affecting the production of specific LPC species (e.g., LPC-DHA, LPC-EPA). Disruption of the LPC profile modifies the Lands’ cycle flux and the profile of LPCs circulating to peripheral tissues. Tissue-level consequences in skeletal muscle and other tissues include: (a) impaired membrane integrity and function; (b) disrupted inflammatory signaling, as LPCs can act as signaling molecules and their fatty acid cargo determines the precursors for pro- vs. anti-inflammatory mediators; and (c) potential impacts on energy metabolism and mitochondrial function. Frailty development emerges as a cumulative consequence of these metabolic, inflammatory, and functional disruptions, exacerbated by the background of inflammaging.
Importantly, this LPC-mediated pathway interacts with direct PFAS toxicity: PFAS-induced oxidative stress can activate PLA2, increasing LPC generation, while LPC itself can promote mitochondrial reactive oxygen species (ROS) production, creating a feed-forward loop[45,46]. Therefore, the direct and indirect pathways are not mutually exclusive but are highly intertwined.
KNOWLEDGE GAPS AND A PATH FORWARD: A TRANSDISCIPLINARY RESEARCH AGENDA
The proposed framework, while plausible, highlights substantial knowledge gaps that require coordinated research efforts. We organize these around three priority questions:
For academia: establishing causality and mechanism
Longitudinal cohort studies are needed to establish the temporal relationship between PFAS exposure (including mixtures and emerging replacements), circulating DHA/EPA and LPC profiles, and incident frailty. Emphasis should be on repeated measurements and adequate power for mediation analyses.
Causal inference methods (e.g., mediation analysis, Mendelian randomization) should be employed as supportive tools to test the proposed pathway, while acknowledging that definitive causality will require triangulation of multiple evidence types.
Experimental studies using age-relevant models (e.g., aged rodents, primary cells from older donors) are crucial to delineate the precise molecular mechanisms by which specific PFAS alter the Lands’ cycle and LPCAT/PLA2 activities, and to determine how PFAS-induced changes in membrane PC composition affect downstream signaling, including SPM production.
For industry and nutrition science: harnessing nutrition for intervention
Pilot or proof-of-concept intervention trials could be considered in well-characterized populations with elevated PFAS exposure or those at high risk of frailty, once sufficient observational evidence establishes consistent mediation. These trials should test whether DHA/EPA supplementation can favorably modulate the LPC profile, reduce inflammation, or improve functional outcomes, but must be designed with appropriate biomarkers and endpoints.
Industry investment in PFAS-free alternatives, remediation technologies, and omega-3-enriched functional foods for vulnerable populations is encouraged, alongside academic partnerships for biomonitoring.
For policymakers: informing integrated risk management
Primary prevention through continued reduction of PFAS emissions and environmental contamination remains the highest priority. Secondary prevention strategies that enhance resilience, such as promoting adequate omega-3 nutrition in exposed populations, may complement exposure reduction but should not substitute for it. Future risk assessments should consider that vulnerable populations (e.g., older adults with poor omega 3 status) may be more susceptible to PFAS-related health effects. This integrated approach aligns with the “One Health” concept, linking environmental contamination, food systems, aging populations, and preventive health.
CONCLUSION
The convergence of an aging global population and ubiquitous exposure to persistent environmental pollutants like PFAS presents a major public health challenge. We propose that the LPC lipid axis, central to membrane phospholipid remodeling and fatty acid trafficking, may be a critical and hitherto overlooked mediator linking PFAS exposure, the protective effects of DHA/EPA, and the pathophysiology of frailty. While LPCs have been recognized as metabolomic biomarkers of frailty, their role as a mechanistic bridge connecting environmental PFAS exposure to frailty pathophysiology through disruption of DHA/EPA-containing phospholipid remodeling represents a novel and testable hypothesis. Elucidating this “PFAS-lipid-frailty nexus” is a scientific imperative that holds promise for informing more effective, integrated strategies for frailty prevention. We end with three testable hypotheses to guide future research:
1. PFAS exposure is associated with altered profiles of DHA/EPA-containing LPC species, and these alterations mediate the association between PFAS and incident frailty.
2. Higher omega-3 status modifies the association between PFAS exposure and frailty risk, with protective effects being most pronounced in individuals with the highest PFAS burdens.
3. PFAS-induced disruption of the Lands’ cycle, reflected in altered LPCAT/PLA2 activity or PC/LPC ratios, is a mechanistic driver of age-related functional decline.
A concerted transdisciplinary effort is now required to translate this framework into actionable knowledge to support a healthier, more sustainable future for all ages.
DECLARATIONS
Acknowledgments
We acknowledge the use of FigDraw 2.0 (https://www.figdraw.com/) for the preparation of the graphical abstract accompanying this manuscript (License ID: UUTUA044b0). Certain icons used in the graphical abstract, including those representing pan, bottled water, food, firefighting, environment, and industry, were sourced from Iconfont (https://www.iconfont.cn/) and incorporated into the graphical design, used in accordance with the applicable licensing terms (CC BY 4.0).
Authors’ contributions
Made substantial contributions to conception and design of the study: Tang, Z.; Ni, J.; Zhu, J.
Performed data analysis and interpretation: Qi, R.; Zhou, Z.; Xu, J.
Performed literature review and data acquisition: Qi, R.; Zhou, Z.; Xu, H.
Wrote the original draft of the manuscript: Qi, R.; Zhou, Z.; Xu, J.
Reviewed and edited the manuscript for important intellectual content: Xu, H.; Ni, J.; Zhu, J.; Tang, Z.
Provided supervision: Ni, J.; Zhu, J.; Tang, Z.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool musely.ai (accessed in 2026) was used solely for the initial drafting of Figure 1. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. The final figure was thoroughly reviewed, edited, and verified by all authors. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
This work was financially supported by the National Natural Science Foundation of China (Grant Nos. 22376039, 21876116).
Conflicts of interest
Zhu, J. is affiliated with Guangdong RangerBio Technologies Co., Ltd., while the other authors have declared no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Fried, L. P.; Tangen, C. M.; Walston, J.; et al.; Cardiovascular Health Study Collaborative Research Group. Frailty in older adults: evidence for a phenotype. J. Gerontol. A. Biol. Sci. Med. Sci. 2001, 56, M146-56.
2. Chen, X.; Giles, J.; Yao, Y.; et al. The path to healthy ageing in China: a Peking University-Lancet Commission. Lancet 2022, 400, 1967-2006.
3. Soysal, P.; Stubbs, B.; Lucato, P.; et al. Inflammation and frailty in the elderly: a systematic review and meta-analysis. Ageing. Res. Rev. 2016, 31, 1-8.
4. Angulo, J.; El Assar, M.; Rodríguez-Mañas, L. Frailty and sarcopenia as the basis for the phenotypic manifestation of chronic diseases in older adults. Mol. Aspects. Med. 2016, 50, 1-32.
5. Xu, Y.; Wang, M.; Chen, D.; Jiang, X.; Xiong, Z. Inflammatory biomarkers in older adults with frailty: a systematic review and meta-analysis of cross-sectional studies. Aging. Clin. Exp. Res. 2022, 34, 971-87.
6. Turkel, I.; Ozerklig, B.; Yılmaz, M.; Ulger, O.; Kubat, G. B.; Tuncer, M. Mitochondrial transplantation as a possible therapeutic option for sarcopenia. J. Mol. Med. 2023, 101, 645-69.
7. Dupont, J.; Dedeyne, L.; Dalle, S.; Koppo, K.; Gielen, E. The role of omega-3 in the prevention and treatment of sarcopenia. Aging. Clin. Exp. Res. 2019, 31, 825-36.
8. Calder, P. C. Omega-3 fatty acids and inflammatory processes: from molecules to man. Biochem. Soc. Trans. 2017, 45, 1105-15.
9. Sunderland, E. M.; Hu, X. C.; Dassuncao, C.; Tokranov, A. K.; Wagner, C. C.; Allen, J. G. A review of the pathways of human exposure to poly- and perfluoroalkyl substances (PFASs) and present understanding of health effects. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 131-47.
10. Evich, M. G.; Davis, M. J. B.; McCord, J. P.; et al. Per- and polyfluoroalkyl substances in the environment. Science 2022, 375, eabg9065.
11. Stanifer, J. W.; Stapleton, H. M.; Souma, T.; Wittmer, A.; Zhao, X.; Boulware, L. E. Perfluorinated chemicals as emerging environmental threats to kidney health: a scoping review. Clin. J. Am. Soc. Nephrol. 2018, 13, 1479-92.
12. Rosen, E. M.; Kotlarz, N.; Knappe, D. R. U.; et al. Drinking water-associated PFAS and fluoroethers and lipid outcomes in the GenX exposure study. Environ. Health. Perspect. 2022, 130, 97002.
13. Tancreda, G.; Campisi, L.; Sarti, M.; Pozzo, L.; Vornoli, A. Perfluoroalkyl substances (PFAS) and lipid metabolism in experimental animal models: a scoping review on the mechanisms behind the induced hepatotoxicity. Curr. Issues. Mol. Biol. 2025, 47, 944.
14. Wielsøe, M.; Long, M.; Ghisari, M.; Bonefeld-Jørgensen, E. C. Perfluoroalkylated substances (PFAS) affect oxidative stress biomarkers in vitro. Chemosphere 2015, 129, 239-45.
15. Liu, B.; Zhu, L.; Wang, M.; Sun, Q. Associations between per- and polyfluoroalkyl substances exposures and blood lipid levels among adults - a meta-analysis. Environ. Health. Perspect. 2023, 131, 56001.
16. Salihovic, S.; Fall, T.; Ganna, A.; et al. Identification of metabolic profiles associated with human exposure to perfluoroalkyl substances. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 196-205.
17. Ma, X.; Cai, D.; Chen, Q.; et al. Hunting metabolic biomarkers for exposure to per- and polyfluoroalkyl substances: a review. Metabolites 2024, 14, 392.
18. Yang, W.; Ling, X.; He, S.; et al. PPARα/ACOX1 as a novel target for hepatic lipid metabolism disorders induced by per- and polyfluoroalkyl substances: an integrated approach. Environ. Int. 2023, 178, 108138.
19. Vanden Heuvel, J. P.; Thompson, J. T.; Frame, S. R.; Gillies, P. J. Differential activation of nuclear receptors by perfluorinated fatty acid analogs and natural fatty acids: a comparison of human, mouse, and rat peroxisome proliferator-activated receptor-alpha, -beta, and -gamma, liver X receptor-beta, and retinoid X receptor-alpha. Toxicol. Sci. 2006, 92, 476-89.
20. Wolf, C. J.; Takacs, M. L.; Schmid, J. E.; Lau, C.; Abbott, B. D. Activation of mouse and human peroxisome proliferator-activated receptor alpha by perfluoroalkyl acids of different functional groups and chain lengths. Toxicol. Sci. 2008, 106, 162-71.
21. Schlezinger, J. J.; Puckett, H.; Oliver, J.; Nielsen, G.; Heiger-Bernays, W.; Webster, T. F. Perfluorooctanoic acid activates multiple nuclear receptor pathways and skews expression of genes regulating cholesterol homeostasis in liver of humanized PPARα mice fed an American diet. Toxicol. Appl. Pharmacol. 2020, 405, 115204.
22. Heindel, J. J.; Blumberg, B.; Cave, M.; et al. Metabolism disrupting chemicals and metabolic disorders. Reprod. Toxicol. 2017, 68, 3-33.
23. Le Magueresse-Battistoni, B.; Vidal, H.; Naville, D. Environmental pollutants and metabolic disorders: the multi-exposure scenario of life. Front. Endocrinol. 2018, 9, 582.
24. Baumert, B. O.; Maretti-Mira, A. C.; Walker, D. I.; et al. Translational framework linking perfluoroheptanoic acid (PFHpA) exposure to metabolic dysfunction associated steatotic liver disease in adolescents. Commun. Med. 2025, 5, 430.
25. Gomis, M. I.; Vestergren, R.; Borg, D.; Cousins, I. T. Comparing the toxic potency in vivo of long-chain perfluoroalkyl acids and fluorinated alternatives. Environ. Int. 2018, 113, 1-9.
26. Wu, B.; Yu, H.; Huang, H.; et al. Efficient biodegradation and detoxification of 6:2 fluorotelomer sulfonic acid by Phanerochaete chrysosporium: insights into enzymatic mechanisms and reduced ecotoxicity. Chem. Eng. J. 2026, 529, 172664.
27. Calder, P. C. Very long-chain n-3 fatty acids and human health: fact, fiction and the future. Proc. Nutr. Soc. 2018, 77, 52-72.
28. Ferri, E.; Marzetti, E.; Calvani, R.; Picca, A.; Cesari, M.; Arosio, B. Role of age-related mitochondrial dysfunction in sarcopenia. Int. J. Mol. Sci. 2020, 21, 5236.
29. Kam, Y.; Winer, L.; Romero, N. Chain length-dependent mitochondrial toxicity of perfluoroalkyl carboxylic acids: insights from Mito Tox Index evaluation. Front. Toxicol. 2025, 7, 1582891.
30. Jia, X.; Liu, W.; Ling, X.; et al. Sex and obesity influence the relationship between perfluoroalkyl substances and lean body mass: NHANES 2011-2018. Heliyon 2024, 10, e35888.
31. Huang, Y.; Wang, C.; Cui, H.; Sun, G.; Qi, X.; Yao, X. Mitochondrial dysfunction in age-related sarcopenia: mechanistic insights, diagnostic advances, and therapeutic prospects. Front. Cell. Dev. Biol. 2025, 13, 1590524.
32. Serhan, C. N.; Levy, B. D. Resolvins in inflammation: emergence of the pro-resolving superfamily of mediators. J. Clin. Invest. 2018, 128, 2657-69.
33. Chiang, N.; Serhan, C. N. Specialized pro-resolving mediator network: an update on production and actions. Essays. Biochem. 2020, 64, 443-62.
34. Bischoff-Ferrari, H. A.; Gängler, S.; Wieczorek, M.; et al. Individual and additive effects of vitamin D, omega-3 and exercise on DNA methylation clocks of biological aging in older adults from the DO-HEALTH trial. Nat. Aging. 2025, 5, 376-85.
35. McGlory, C.; Calder, P. C.; Nunes, E. A. The influence of omega-3 fatty acids on skeletal muscle protein turnover in health, disuse, and disease. Front. Nutr. 2019, 6, 144.
36. Kim, J.; Kim, M.; Won, C. W.; Park, Y. Erythrocyte levels of n-3 polyunsaturated fatty acids and incidence of frailty after a 6-year follow-up: the Korean frailty and aging cohort study. Front. Nutr. 2025, 12, 1569832.
37. Gagesch, M.; Wieczorek, M.; Vellas, B.; et al. Effects of vitamin D, omega-3 fatty acids and a home exercise program on prevention of pre-frailty in older adults: the DO-HEALTH randomized clinical trial. J. Frailty. Aging. 2023, 12, 71-7.
38. Hashidate-Yoshida, T.; Harayama, T.; Hishikawa, D.; et al. Fatty acid remodeling by LPCAT3 enriches arachidonate in phospholipid membranes and regulates triglyceride transport. Elife 2015, 4, e06328.
39. Domingo, J. L. Health risks of dietary exposure to perfluorinated compounds. Environ. Int. 2012, 40, 187-95.
40. O’Donnell, V. B. New appreciation for an old pathway: the lands cycle moves into new arenas in health and disease. Biochem. Soc. Trans. 2022, 50, 1-11.
41. Meng, L.; Yang, R.; Wang, D.; et al. Specific lysophosphatidylcholine and acylcarnitine related to sarcopenia and its components in older men. BMC. Geriatr. 2022, 22, 249.
42. Meng, L.; Shi, H.; Wang, D. G.; et al. Specific metabolites involved in antioxidation and mitochondrial function are correlated with frailty in elderly men. Front. Med. 2022, 9, 816045.
43. Takagi, S.; Maeda, K.; Takemura, M.; et al. Low plasma lysophosphatidylcholine levels are associated with reduced handgrip strength in older outpatients. Geriatr. Gerontol. Int. 2026, 26, e70281.
44. Sevastou, I.; Kaffe, E.; Mouratis, M. A.; Aidinis, V. Lysoglycerophospholipids in chronic inflammatory disorders: the PLA2/LPC and ATX/LPA axes. Biochim. Biophys. Acta. 2013, 1831, 42-60.
45. Zou, Y.; Kim, D. H.; Jung, K. J.; et al. Lysophosphatidylcholine enhances oxidative stress via the 5-lipoxygenase pathway in rat aorta during aging. Rejuvenation. Res. 2009, 12, 15-24.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].