


Ranking the gut microbiome for cardiometabolic health: the good, the bad and the ugly
 0
0 Rethinking the microbial “good”, “bad”, and “ugly”
Cardiometabolic diseases are shaped not only by diet, dyslipidemia, and chronic low-grade inflammation, but also by the gut microbiome, a modifiable component of the same biological axis. Yet one persistent obstacle in the field has been the difficulty of defining what a “good” or “bad” microbiome actually is. It is just like the classic film The Good, the Bad and the Ugly: on the path to gold, the good, the bad, and the ugly—much like beneficial and harmful bacteria—compete for their contribution to the host's metabolism. This conceptual framework is illustrated in Figure 1. Interindividual variation in host genetics, medication exposure, dietary habits, geography, and lifestyle has made any universal microbial template elusive. In this context, the study by Asnicar et al.[1] is notable less for proposing a single ideal microbiome than for reframing the question itself: which microbial species can, across cohorts and populations, consistently track with a more favorable cardiometabolic state?
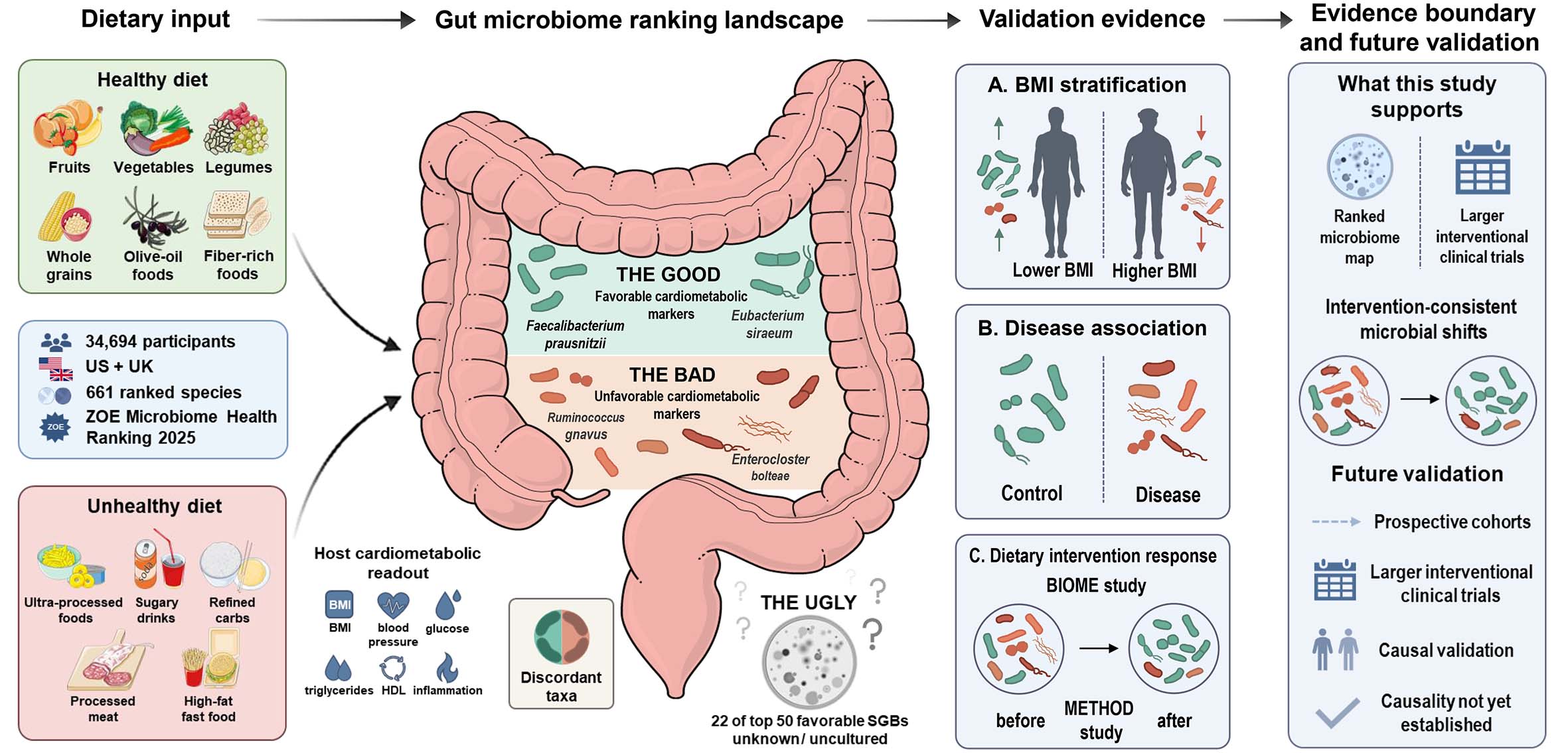
Figure 1. Conceptual summary of the gut microbiome ranking framework for cardiometabolic health. In the original study, 661 gut microbial species were ranked in 34,694 participants from the US and UK according to their associations with dietary patterns and cardiometabolic markers. This schematic summarizes the proposed framework, highlighting favorably ranked taxa (“the good”), unfavorably ranked taxa (“the bad”), and unknown or uncultured candidates (“the ugly”). The right-hand validation section summarizes three lines of supporting evidence: (A) BMI stratification, in which lower BMI is associated with a greater representation of favorably ranked microbes and higher BMI with a greater representation of unfavorably ranked microbes; (B) disease association, in which control or healthier states show relatively more favorable microbial signatures, whereas disease states show relatively more unfavorable signatures; and (C) dietary intervention response, in which the BIOME and METHOD intervention studies suggest that diet-related interventions can shift the gut microbiome toward a more favorable configuration. Together, these findings support the reproducibility and potential actionability of the microbiome ranking landscape. Further prospective studies, larger interventional trials, and causal validation will likely be needed. Selected graphical elements were adapted from Servier Medical Art (https://smart.servier.com), licensed under CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/), and from Bioicons elements identified as public domain/no restrictions in the accompanying attribution/license table. Graphical elements were modified in color, size, orientation, and layout.
A major strength of this study lies in its ability to translate a highly complex and deeply individualized ecological system into a framework that is comparable, testable, and open to biological validation. Rather than merely listing differentially abundant taxa, the authors integrated partial Spearman correlations between 661 non-rare microbial species and 37 intermediate cardiometabolic markers, thereby constructing a continuous gradient from more favorable to less favorable taxa. Importantly, this ranking was not confined to the internal dataset, but also showed consistent associations with BMI [Figure 1A] and disease status in external public metagenomic cohorts [Figure 1B].
The study also goes beyond showing which microbes differ by offering an initial indication of how they may shift under intervention. In the biotics influence on microbiome ecosystem (BIOME) and measuring efficacy through outcomes of diet (METHOD) nutritional intervention studies, favorably ranked taxa tended to increase, whereas unfavorably ranked taxa tended to decrease [Figure 1C]. For cardiovascular research, this observation is especially relevant because it suggests that these species may represent more than passive correlates of host state. They may instead capture an intermediate layer through which dietary intervention is transmitted to microbial remodeling and, ultimately, to host benefit. This reflects the important role of so-called "the good" and "the bad" bacteria in transmitting dietary signals, thereby regulating the progression of metabolic cardiovascular disease.
From taxonomic ranking to mechanistic interpretation
From a mechanistic perspective, however, a linear ranking of individual species should not be mistaken for a complete description of gut ecology. The gut microbiome is fundamentally a nonlinear, context-dependent metabolic network shaped by competition, mutualism, and cross-feeding. Asnicar et al. also identified so-called discordant taxa, namely species associated with less favorable dietary patterns yet linked to more favorable cardiometabolic markers[1]. This point is more than theoretical and may be termed "the good and the bad" referring to the opposite roles that the same species can exhibit under different conditions. A recent atrial fibrillation study, for example, suggested that Ruminococcus gnavus, a species not typically viewed as uniformly beneficial, may attenuate atrial fibrosis and susceptibility to atrial fibrillation through the production of isovaleric acid (IVA)[2]. Such observations underscore that the “health weight” assigned to an individual species does not necessarily reflect its isolated function. More often, it reflects the ecological and metabolic position that species occupies within a broader community network. For that reason, the ZOE ranking is best understood not as a definitive table of single-species pathogenic or protective effects, but as a prioritized map of candidates for mechanistic follow-up.
For cardiovascular research, the real value of this study may therefore lie in forcing the gut microbiota-metabolite-vascular axis to be interpreted simultaneously at ecological, metabolic, and pathological levels.
At the ecological level, the more relevant question is not whether one isolated “beneficial bacterium” increases, but whether a metabolically cooperative consortium is activated as a whole. Judging from the intervention results in this study, what increased was not a single “star microbe”, but rather something closer to a coordinated functional network. Bifidobacterium adolescentis, for example, is more likely to function as an upstream fiber utilizer, first producing intermediate metabolites such as lactate and acetate, which are then further converted by downstream taxa including Anaerobutyricum hallii, Agathobaculum butyriciproducens, and Roseburia hominis, thereby promoting butyrate production. These shifts are therefore better understood as activation of a metabolic relay network favorable for short-chain fatty acid generation, rather than as the isolated effect of one organism[3]. For cardiovascular research, this distinction matters, because it suggests that the truly important question for future work may not be which single taxon is “best”, but which microbial consortia and metabolic cooperativity most effectively influence host inflammatory tone and lipid metabolism.
At the metabolic level, it is no longer sufficient to say simply that “SCFAs are anti-inflammatory.” A more precise formulation is that short-chain fatty acids, especially butyrate, may contribute to cardiometabolic protection through at least three relatively well-defined routes. First, butyrate supports colonocyte energy supply and barrier integrity, thereby reducing the translocation of endotoxin into the circulation that accompanies increased intestinal permeability. Second, SCFAs can influence Treg homeostasis, monocyte-macrophage inflammatory responses, and the host immune threshold through signaling pathways involving FFAR2 (free fatty acid receptor 2), FFAR3 (free fatty acid receptor 3), and GPR109A/HCAR2 (G protein-coupled receptor 109A/hydroxycarboxylic acid receptor 2), as well as through epigenetic regulation. Third, these effects are unlikely to remain confined to the intestinal compartment; rather, they may constitute an upstream condition through which diet-related microbial signals shape distal metabolic organs and the vascular system[4,5]. In other words, SCFAs should be viewed not merely as beneficial metabolites, but as an important intermediate layer linking dietary structure, gut ecology, and host inflammatory background. A necessary note of restraint, however, is that the Nature study by Asnicar et al. did not directly measure SCFA flux and did not demonstrate that the highly ranked taxa necessarily act through any one specific metabolite pathway[1]. The mechanistic extension proposed here is therefore best understood as a reasoned bridge between the present findings and prior literature, rather than as a direct answer already delivered by this study.
At the pathological level, the key question is no longer whether SCFAs are beneficial in general, but whether they can influence the cellular programs most proximal to cardiometabolic injury. For atherosclerosis in particular, this means processes within the plaque itself, including macrophage-driven lipid uptake and cholesterol efflux, foam cell formation, inflammatory activation, necrotic core expansion, and plaque instability. Experimental studies suggest that butyrate can attenuate plaque inflammation, promote ABCA1 (ATP-binding cassette subfamily A member 1)-dependent macrophage cholesterol efflux, and modulate macrophage polarization[6-8]. The real value of highly ranked taxa, therefore, may lie less in their association with healthier diets per se than in whether the metabolic networks they represent can alter these disease-relevant processes. That conclusion cannot yet be drawn directly from the data of Asnicar et al.[1]. But that is precisely why the study matters: rather than simply sorting microbes into healthier and harmful groups, or so-called “the good and the bad” species, it provides cardiometabolic diseases (CMD) research, and atherosclerosis research in particular, with a structured species-level map for asking a more translational question: which microbial assemblies, through which metabolites, influence which vascular and cardiometabolic pathological processes?
Toward cardiovascular microbiome translation
For cardiovascular research, perhaps the most consequential contribution of this study is that it has sharply clarified where the field’s real mechanistic gap lies. Most notably, it brings microbial “dark matter” to the forefront: Asnicar et al. found that 22 of the 50 most favorably ranked species-level genome bins remain unknown and uncultured, existing only as metagenome-assembled genomes[1]. These are referred to as “the ugly”, indicating that their precise roles have yet to be determined. We can assign them health-associated weights across tens of thousands of individuals, yet still cannot culture them, colonize them in isolation, or test them causally in standardized models. The challenge now is not simply to refine the ranking, but to turn these high-priority yet inaccessible microbes into experimentally tractable functional units.
Sex differences should also be considered when translating such microbial rankings into cardiovascular mechanisms. In the original study, sex was included as a covariate in key analyses, including species-ranking construction and BMI- or disease-related validation[1]. However, adjustment for sex does not answer whether the same ranked taxa have equivalent biological meanings in women and men. Given evidence that sex hormones can shape gut microbial ecology and host metabolic phenotypes, future studies should test sex-stratified microbiome rankings and sex-by-microbiome interactions, particularly across menopausal transition and other sex-specific cardiometabolic risk states[9,10].
Overall, the work of Asnicar et al. provides CMD research, and atherosclerosis research in particular, with an unprecedentedly structured species-level map for navigation[1]. Its academic value lies in compressing vast and noisy association signals into a priority system that can be dissected biologically, step by step. For cardiovascular investigators, the next step worth pursuing is to align this ranking map more precisely with vascular wall pathology, and to determine which highly ranked taxa, through which combinations of metabolites, influence gut barrier function, reset immune thresholds, promote macrophage cholesterol efflux, and ultimately alter plaque inflammation and stability. Only when the “good” are mechanistically defined, the “bad” functionally understood, and the “ugly” rendered experimentally accessible, can the gut microbiome move from a diet-associated biomarker to an actionable target in cardiovascular precision medicine. That transition may define one of the most consequential next phases of cardiovascular microbiome research.
DECLARATIONS
Authors’ contribution
Drafted the manuscript, performed the literature review, prepared Figure 1, and revised the manuscript: Guo M
Conceived the commentary, provided intellectual guidance, and critically revised the manuscript: Li T
Supervised the work, provided critical academic input, and critically revised the manuscript: Yuan Z
All authors read and approved the final version of the manuscript.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool Gemini 2.5 Pro (Google; version gemini-2.5-pro, released 2025-06-17) was used solely for language editing. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
This work was supported by the following funding sources: National Key R&D Program of China Grant (2021YFA0805400, 2024YFA1307004); The Project of National Science Foundation of China (No. 82370458, 82430019, 82570531); Natural Science Basic Research Program of Shaanxi Province (2025JC-YBQN-1229).
Conflicts of interest
Li T is the Guest Editor of the special issue "Gut Microbiota and Metabolites in Cardiovascular Aging and Disease" in The Journal of Cardiovascular Aging, and also is a member of the journal’s Editorial Board. Li T was not involved in any steps of the editorial process for this manuscript, including reviewer selection, manuscript handling, or decision-making, The other authors declare that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Asnicar F, Manghi P, Fackelmann G, et al. Gut micro-organisms associated with health, nutrition and dietary interventions. Nature. 2025;650:450-8.
2. Ding N, Wu H, Hua Y, et al. Gut microbiota-derived isovaleric acid alleviates atrial fibrillation by suppressing GSDME-dependent pyroptosis. Cell Metab. 2026;38:370-87.e10.
3. Zhao L, Zhang F, Ding X, et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science. 2018;359:1151-6.
4. Koh A, De Vadder F, Kovatcheva-Datchary P, Bäckhed F. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell. 2016;165:1332-45.
5. Macia L, Tan J, Vieira AT, et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat Commun. 2015;6:6734.
6. Fusco W, Adolph T, Cammarota G, Gasbarrini A, Ianiro G, Tilg H. Gut microbiota and atherosclerosis. Gut. 2026;75:1067-77.
7. Valles-Colomer M, Menni C, Berry SE, Valdes AM, Spector TD, Segata N. Cardiometabolic health, diet and the gut microbiome: a meta-omics perspective. Nat Med. 2023;29:551-61.
8. Marques FZ, Mackay CR, Kaye DM. Beyond gut feelings: how the gut microbiota regulates blood pressure. Nat Rev Cardiol. 2017;15:20-32.
9. Markle JGM, Frank DN, Mortin-Toth S, et al. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science. 2013;339:1084-8.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].