


Role of mechanical forces and skin tension in scar pathophysiology
 0
0 
Abstract
Scar formation is the natural consequence of wound healing, yet in many cases it leads to hypertrophic scars or keloids that impair function, create aesthetic concerns, and diminish quality of life. While biochemical signaling pathways in wound repair have been well characterized, mechanical forces are now recognized as equally crucial regulators of cutaneous fibrosis. This narrative review utilized a structured literature search of PubMed and Google Scholar databases to evaluate experimental, translational, and clinical evidence regarding the role of mechanical tension and mechanotransduction in wound healing and pathological scar formation. Relevant studies investigating biomechanical signaling pathways, fibroblast activation, extracellular matrix remodeling, and therapeutic tension-modulating strategies were identified through title, abstract, and full-text review. We outline the four phases of wound repair and highlight how aberrant mechanical inputs promote persistent inflammation, fibroblast overactivation, excessive extracellular matrix deposition, and impaired collagen remodeling. Key mechanotransduction pathways, including focal adhesion kinase (FAK)-extracellular signal-regulated kinase (ERK)-monocyte chemoattractant protein-1 (MCP-1), transforming growth factor-beta (TGF-β)/Smad, platelet-derived growth factor (PDGF)-PI3K/AKT and mitogen-activated protein kinase (MAPK), Yes-associated protein (YAP)/transcriptional coactivator with PDZ-binding motif (TAZ), and Wnt/β-catenin, are discussed with emphasis on their roles in myofibroblast differentiation and fibrosis. Translational advances including stress-shielding devices, force-modulating closure techniques, and topical or intradermal inhibitors of FAK or YAP demonstrate how targeting mechanical pathways can significantly reduce scar formation in animal models and preclinical studies. As mechanical biology continues to intersect with regenerative medicine, therapeutics that modulate tension or interrupt profibrotic mechanotransduction hold promise for shifting cutaneous repair from a fibrotic toward a regenerative, scar-minimizing paradigm.
Keywords
INTRODUCTION
Wound healing is a complex and dynamic biological process through which the body restores tissue integrity following an insult or injury, with scar formation as the natural endpoint. Although scarring is vital for quickly closing wounds and re-establishing the skin’s protective barrier, it often leads to fibrotic tissue that lacks the integrity and functionality of healthy skin. This can result in functional limitations with restricted mobility of the tissue, aesthetic issues, and emotional distress for patients[1]. Beyond their physical manifestations, abnormal scars can impose substantial psychosocial burden, negatively affecting self-esteem, body image, interpersonal relationships, and social perception. Patients frequently report emotional distress, social anxiety, and reduced quality of life related to the visibility and chronicity of these lesions. Given the high prevalence of pathological scarring and the limitations of existing therapies, understanding the molecular and biomechanical mechanisms underlying fibrosis remains essential for the development of more effective preventative and therapeutic strategies.
Following a traumatic insult that disrupts skin integrity, the normal process of cutaneous wound repair engages a myriad of cell types in a strictly coordinated fashion. This process involves four primary phases: hemostasis, inflammation, proliferation, and remodeling. Each step is regulated by a cascade of cellular and molecular events and mobilizes various cell types, including platelets, immune cells, fibroblasts, macrophages, and keratinocytes, to repair damaged tissue and restore skin function[1-4]. Abnormal scar development, such as hypertrophic scars and keloids, is marked by prolonged inflammation, overactive fibroblasts, excessive collagen buildup, and disrupted remodeling of the tissue matrix[5]. Factors such as mechanical stress and skin tension are known to significantly influence these changes[5]. Gaining deeper insight into the biology of scar formation is essential for designing strategies that reduce or prevent problematic scarring and enhance healing outcomes, particularly given its significant psychosocial impact, including effects on self-esteem, body image, and social perception, which can substantially impair patients’ quality of life. This review provides a translational overview of current evidence surrounding mechanical tension, mechanotransduction pathways, and emerging tension-modulating therapies in hypertrophic and keloid scar formation. By integrating preclinical and early clinical evidence, this manuscript highlights mechanical signaling as both a key driver of fibrosis and a promising therapeutic target for scar prevention.
Throughout this review, several biomechanical terms are used that warrant clarification. Mechanical forces is the broadest term and encompasses all physical stimuli acting on tissues, including tension, compression, shear stress, and strain. Skin tension refers to the intrinsic resting mechanical stress present within the skin as a result of collagen fiber organization, elastin architecture, and regional biomechanical properties, often reflected by Langer’s lines. In contrast, mechanical tension refers to externally applied or dynamically generated tensile forces acting on tissues during movement, wound contraction, surgical closure, or tissue expansion. Although multiple forms of mechanical force can influence cellular behavior, the majority of evidence linking biomechanics to pathological scar formation has focused on tensile loading and tension-mediated mechanotransduction. Therefore, the term “tension” is used throughout this review to describe the predominant mechanical stimulus implicated in hypertrophic scar and keloid development.
METHODS
This narrative review was conducted through a comprehensive literature search of PubMed and Google Scholar databases to evaluate current evidence regarding the roles of mechanical forces and mechanotransduction in wound healing and pathological scar formation. The literature search was performed between January 2025 and March 2025 and mostly focused on reviews from the last 5 years, however, some older data that strongly relates to the review were also included.
Search terms included combinations of keywords and Medical Subject Headings (MeSH) terms such as “scar formation”, “wound healing”, “mechanical forces”, “mechanotransduction”, “skin tension”, “hypertrophic scar”, “keloid”, “fibrosis”, “focal adhesion kinase”, “YAP/TAZ”, “transforming growth factor-beta”, and “extracellular matrix”. Boolean operators (AND/OR) were utilized to refine the search strategy and identify studies relevant to mechanobiology and scar pathophysiology.
Articles were screened initially by title and abstract review, followed by full-text assessment when appropriate. Studies were included if they investigated the relationship between biomechanical forces and wound healing, hypertrophic scar formation, keloid development, fibroblast mechanotransduction, extracellular matrix (ECM) remodeling, or therapeutic strategies targeting mechanical signaling pathways. Both experimental and clinical studies were considered eligible for inclusion. Foundational mechanobiology studies and relevant review articles were additionally included to provide historical context and support conceptual discussion.
Studies were excluded if they were unrelated to cutaneous wound healing, focused solely on non-skin fibrotic diseases, lacked mechanobiological relevance, were non-English publications, or consisted of conference abstracts without sufficient methodological detail. Preference was given to peer-reviewed studies with clear experimental methodology and translational relevance.
The majority of included studies consisted of preclinical investigations utilizing murine models, porcine models, in vitro fibroblast studies, and mechanobiological experiments evaluating tensile stress and fibrosis-related signaling pathways. Due to the limited availability of high-quality randomized human clinical trials in this field, human evidence included observational studies, early clinical trials, and translational investigations involving scar reduction therapies and tension-offloading devices.
Because of heterogeneity in study design, experimental models, and reported outcomes, a quantitative meta-analysis was not performed. Instead, findings were synthesized narratively with emphasis placed on mechanotransduction pathways, biomechanical regulation of fibrosis, and translational therapeutic applications.
ETIOLOGY AND EPIDEMIOLOGY OF PATHOLOGIC SCARS
Pathological scar formation represents a significant burden to the global healthcare system, resulting in repeated clinical interventions, long-term patient effects, and billions of dollars spent yearly[6]. Two of the most common types of pathological scars are hypertrophic scarring (HTS) and keloid scars. While similar in origin and development, several key differences exist, including scar extent, causality, and genetic predisposition[7]. Increasing evidence suggests that mechanical tension and biomechanical forces play an important role in the development and progression of both scar types. While HTS formation is often observed in burn victims (who suffer from HTS rates up to 70% when affected by deep burns), keloid scars are more commonly observed in adolescent populations, which may imply a link to higher hormone levels and increased skin tension during periods of rapid growth[7,8]. Additionally, the highest rates of keloid scar formation are seen in individuals living in equatorial regions, especially in African, Asian, and Latin American countries[9]. While it is unclear to what degree this increased rate is due to increased sun and UV exposure in these areas, unequal access to treatments, genetics, or differences in skin biomechanics, these remain established epidemiologic factors[7].
Keloids are a form of abnormal or pathological scarring that arise when the wound-healing pathway becomes dysregulated. Specifically, keloids are a benign fibroproliferative disorder in which excessive amounts of collagen are produced by cells during the wound-healing process, resulting in an enlarged scar that often extends beyond the original borders of the wound[10,11]. The keloid scar is characterized by an exaggerated macroscopic appearance as well as histology revealing a random meshwork of collagen bundles, which differs from the linear collagen arrangement of regular scarring[12,13]. Keloids also frequently occur in regions of elevated cutaneous tension, including the chest, shoulders, upper back, and joints, supporting the theory that mechanical stress contributes to their pathogenesis. The exact pathogenesis of keloid scars remains unclear, with various theories proposed and researched, including sustained fibroblast activation, disordered epithelial-mesenchymal transition, excessive angiogenesis, and aberrant mechanotransduction signaling[14-16]. Genetics, age, and gender predispositions towards keloid formation have been identified as well[17]. Keloid scars can be quite burdensome for those affected, as they are both aesthetically inferior and often uncomfortable or painful. Keloids are a challenging clinical obstacle to overcome for physicians, as they are often resistant to therapeutic options and have high rates of recurrence after being treated[18].
Similarly, a hypertrophic scar arises when the normal wound healing process becomes exaggerated and an excess amount of collagen is deposited, leading to a raised and thickened scar[19]. Unlike a keloid, a hypertrophic scar remains within the confines of the original wound boundary and does not spread into surrounding healthy skin[20]. Hypertrophic scars are especially common in wounds subjected to prolonged tension, repetitive motion, or delayed healing, such as burns over joints or highly mobile regions of the body. The result of such scarring is often cosmetically inferior and results in patient dissatisfaction, often driving individuals to seek various forms of treatment to improve the appearance of their scars. Various interventions are currently in use to combat HTS, including surgical scar revision, corticosteroid injections, topical treatments, laser therapy, and emerging tension-reducing therapies[21-23].
Certain anatomic regions are more prone to hypertrophic scar and keloid formation, which may indicate the mechanism of scar formation. Areas like the sternum, shoulders, upper back, and joints develop scar tissue more than other parts of the body. This is of particular interest because these locations are exposed to elevated mechanical tension, repetitive stretching, and greater levels of mobility, all of which may contribute to prolonged inflammatory signaling and abnormal collagen deposition during healing.
While the exact mechanisms underlying pathological scar formation remain incompletely understood, the normal wound healing pathway is likely affected by both intrinsic cellular abnormalities and extrinsic mechanical forces. By investigating the role of various cellular signaling pathways and how they can be manipulated, as well as the external forces that act upon the overall process, clinicians and scientists may find actionable targets to reduce or entirely prevent pathological scarring. Success in this field, whether partial or complete, will reduce the impact of these prominent scars on patients and remove the burden of treatment and revision from healthcare professionals. In this manuscript, we discuss the role of mechanical forces in wound healing and scar formation as well as current and developing therapeutic options.
STAGES OF WOUND HEALING
Wound healing proceeds through four major stages: hemostasis, inflammation, proliferation, and maturation/remodeling. Although each phase contributes to tissue repair, mechanical tension primarily exerts its effects during the proliferative and remodeling stages, where it influences fibroblast activity, ECM deposition, wound contraction, and scar formation.
The proliferative phase of wound healing is strongly influenced by mechanical tension and mechanotransduction during granulation tissue formation, fibroblast activation, and wound contraction[1-4]. During this stage, growth factors such as transforming growth factor-beta (TGF-β) and platelet-derived growth factor (PDGF) recruit and activate fibroblasts within the wound bed[24-26]. Fibroblasts proliferate and synthesize components of the provisional ECM, including type III collagen, proteoglycans, and glycosaminoglycans, which provide structural support and a scaffold for cellular attachment and migration[27,28]. As mechanical forces accumulate within the healing tissue, fibroblasts sense and respond to matrix stiffness and tensile stress through integrins and associated cytoskeletal signaling pathways. These biomechanical cues contribute to fibroblast differentiation into contractile myofibroblasts[29-32].
The activation of signaling pathways such as TGF-β/Smad, PDGF/AKT, and PDGF/mitogen-activated protein kinase (MAPK) promotes myofibroblast differentiation, characterized by increased expression of α-smooth muscle actin (α-SMA) and enhanced synthesis of ECM proteins including type I and III collagen, fibronectin, and hyaluronic acid[27,29,31,32]. Myofibroblasts generate contractile forces through actin-myosin interactions that are mechanically transmitted to the ECM via focal adhesions. These forces facilitate wound contraction, physically reducing wound size and drawing wound edges together[33-35]. Mechanical tension further reinforces myofibroblast persistence and activity, creating a feedback loop in which increased tissue stiffness promotes continued cellular contraction and ECM deposition.
Mechanical tension also plays a critical role during the maturation, or remodeling, phase of wound healing[36-38]. During this stage, the initially deposited type III collagen is gradually degraded by matrix metalloproteinases (MMPs) and replaced with stronger, more organized type I collagen[36,38]. Importantly, collagen fibers align along lines of mechanical stress, thereby improving the tensile strength and structural integrity of the repaired tissue. As remodeling progresses, fibroblast density and vascularity decrease, and the wound transitions into a less cellular and more mechanically stable scar[36,38,39].
The balance of mechanical signaling during remodeling is essential for normal scar formation. Under physiologic conditions, scar tissue matures into a relatively flat and pliable structure composed of organized collagen bundles[40,41]. However, excessive or prolonged mechanical tension can dysregulate mechanotransductive signaling pathways, contributing to pathological scarring such as hypertrophic scars and keloids[19,23,42,43]. Elevated tensile forces promote sustained fibroblast and myofibroblast activation, excessive collagen synthesis, and reduced ECM degradation, ultimately leading to abnormal scar accumulation and tissue stiffness.
MECHANICAL FORCES
As previously mentioned, several physiological and biomechanical factors drive the normal process of wound healing and scar formation. Among these, mechanical forces and tension have long been recognized as crucial contributors, with increasing research over recent decades elucidating their role in scar biology.
Langer’s lines are a historic representation of the natural patterns of tension in human skin, arising from the orientation of collagen fibers and fibroblast activity. Surgical incisions aligned with these lines are exposed to less stress and are traditionally believed to heal with less obtrusive scars, while those placed perpendicular to the lines are more likely to widen and form prominent scars[44]. Karl Langer, who first described these tension lines, mapped them by puncturing cadaver skin and noting how circular wounds elongated into clefts that reflected the underlying collagen alignment. His topographical findings established that skin mechanics are closely tied to dermal structure, with collagen organization dictating how skin responds to injury and surgical manipulation. These early insights laid the foundation for the modern surgical approach to creating incisions, where respecting Langer’s lines can reduce scar formation and wound complications. Building on this, surgeon Dr. Robert Wray later quantified the relationship between wound closure force and scar width, showing that both intrinsic skin tension and external mechanical forces play crucial roles in healing and scar development[45]. Using a porcine model, outcomes of wound healing under artificially applied tension, normal tension, and tension-offloading were recently compared. Remarkably, tension-offloading reduced histologic scar area six-fold compared to controls and nine-fold compared to the applied tension group[46]. This was further investigated in human trials, which showed that stress shielding resulted in decreased scar formation when compared to controls. Following these small human clinical studies, this idea of offloading mechanical tension has been utilized in surgical practice through the use of wound support devices such as steri-strips, as well as through specific techniques in plastic and reconstructive surgery such as the Z-plasty incision[47,48].
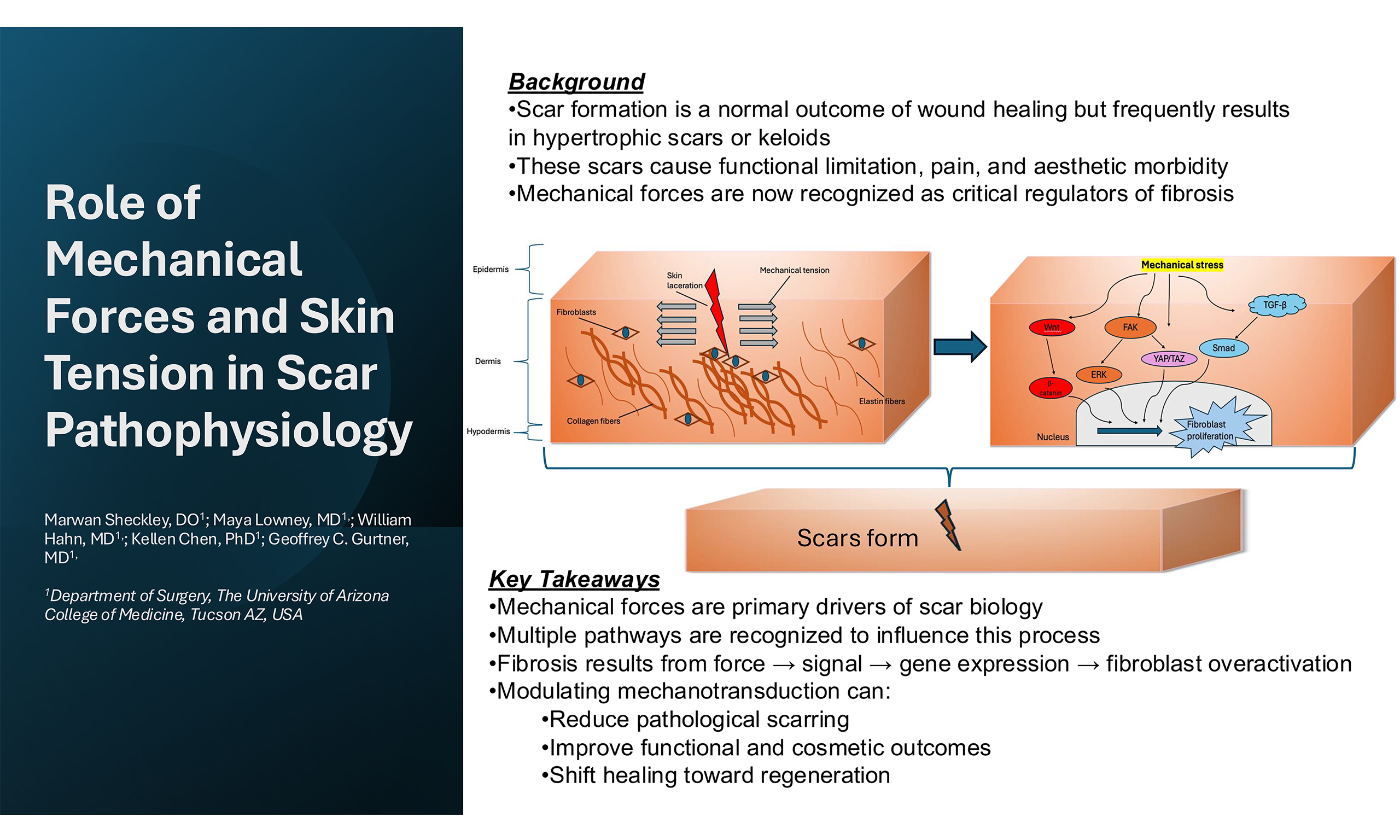
The influence of mechanical forces on scar formation is a result of the aggregation of multiple pathways, not any single signaling cascade [Figure 1]. Through the conversion of physical tension into cellular signals, these pathways will activate various cell types that contribute to the formation of a profibrotic state. They collectively regulate fibroblast differentiation, collagen synthesis, and ECM remodeling during scar development.
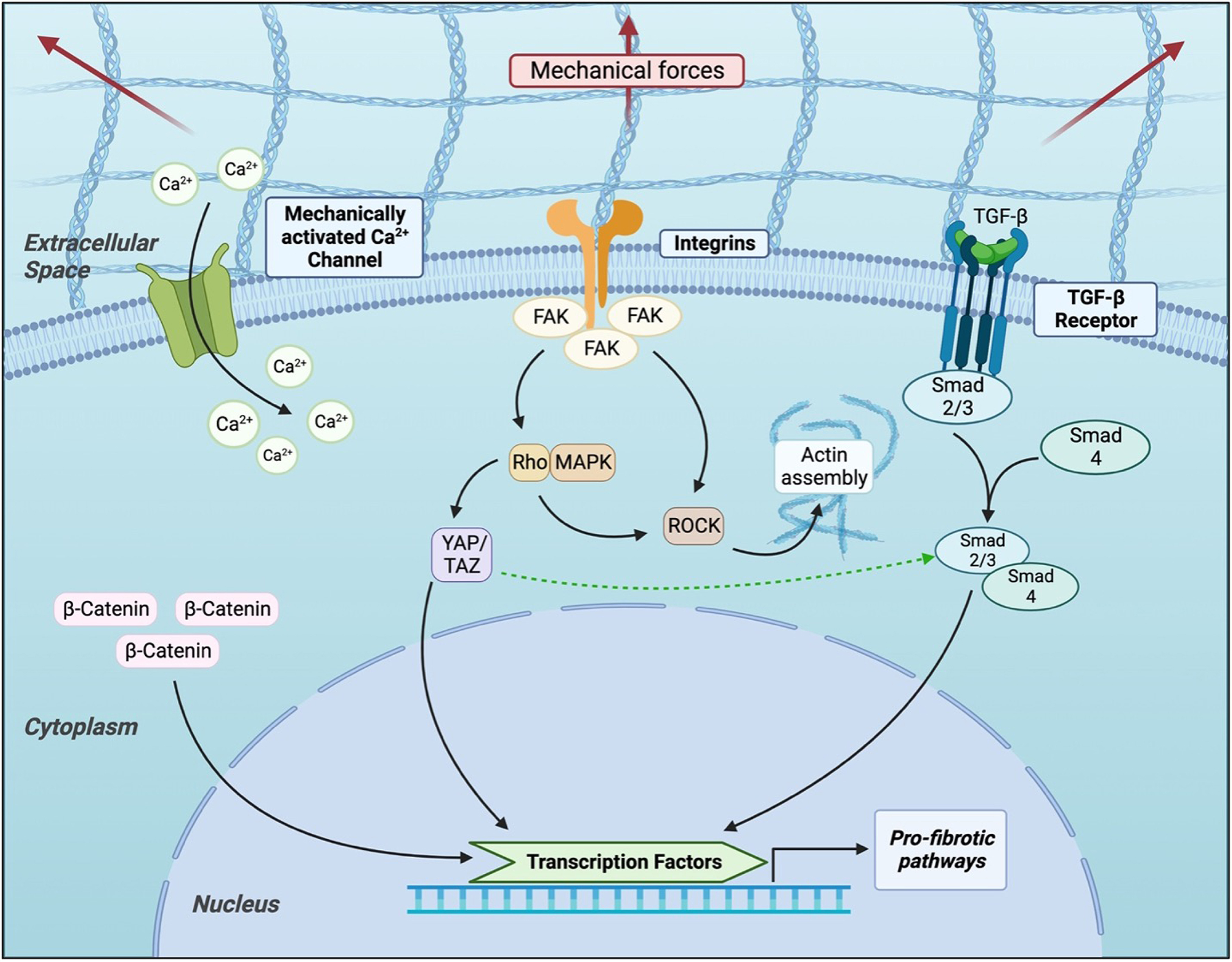
Figure 1. Cellular signaling pathways in mechanotransduction in scar formation. Adapted from Berry et al., Frontiers in Surgery, 2023[49], licensed under CC BY 4.0. FAK: Focal adhesion kinase; MAPK: mitogen-activated protein kinase; ROCK: Rho-associated coiled-coil containing protein kinase; YAP: Yes-associated protein; TAZ: transcriptional coactivator with PDZ-binding motif; TGF-β: transforming growth factor-beta; Smad: suppressor of mothers against decapentaplegic homolog.
At the cellular level, cells are embedded within a protein-dense structure called the ECM, which offers mechanical support as well as facilitates cell-to-cell communication. The ECM is integral to wound healing, playing a central role in modulating signaling pathways and regulating key cellular behaviors such as proliferation, migration, and survival. Although composed of many different proteins, collagen and elastin are among the most prevalent components of the ECM[50]. Type I and type III collagen are arranged in an irregular, basket-weave-like architecture. This disordered structure imparts tensile strength and rigidity to the skin, enhancing its ability to resist permanent deformation and tearing. In contrast, elastin fibers are extensively cross-linked and provide the skin with its elastic recoil, allowing it to stretch and return to its original shape after mechanical stress is removed. Together, collagen and elastin form intertwined fibrous networks that underlie the characteristic “J-shaped” stress-strain response of skin when subjected to uniaxial tension[51,52]. This response includes three distinct phases: (i) an initial low-stiffness region where elastin fibers align with increasing load; (ii) a linear stiffening phase as collagen fibers straighten and bear load; and (iii) a high-strain region culminating in tissue deformation, rupture, and scarring[53]. These fibrous proteins interact with cells primarily through integrins, which are transmembrane heterodimeric receptors that serve as conduits for bidirectional signal transmission between the cell’s exterior and its internal environment. Integrins physically link the ECM to the actin cytoskeleton inside the cell and are central to the structure and function of Focal Adhesions[54-56].
MECHANOTRANSDUCTION AND OTHER SIGNALING PATHWAYS IMPORTANT TO FIBROSIS
Focal adhesion kinase
Focal Adhesions are multiprotein complex structures that have garnered significant attention in the study of mechanotransduction due to their ability to convert mechanical stimuli into biochemical signals. When external forces are applied to the ECM, Focal Adhesions translate these mechanical inputs into intracellular responses, activating signaling cascades that promote inflammatory cell recruitment, keratinocyte movement, angiogenesis, elevated collagen production, and profibrotic gene expression via Yes-associated protein (YAP) [Figure 2]. All of these processes combine to contribute to scar formation. In a genome-wide analysis of murine cells subjected to mechanical force, focal adhesion kinase (FAK) was identified to be a pivotal mechanotransduction regulator that orchestrates fibrotic signaling via the extracellular signal-regulated kinase (ERK) pathway and promotes monocyte chemoattractant protein-1 (MCP-1) expression. FAK itself is a cytoplasmic tyrosine kinase that serves as an important mediator in several pathways[57]. Increased mechanical tension across a wound has been shown to markedly elevate FAK activation (expressed via autophosphorylation at Tyr-397) and resultant downstream signaling through ERK to affect collagen production[58]. The effects of FAK modulation are reflected both at a molecular and macrocosm level, with outcomes such as changes in functional biomechanical properties including tissue elasticity and compliance[59].
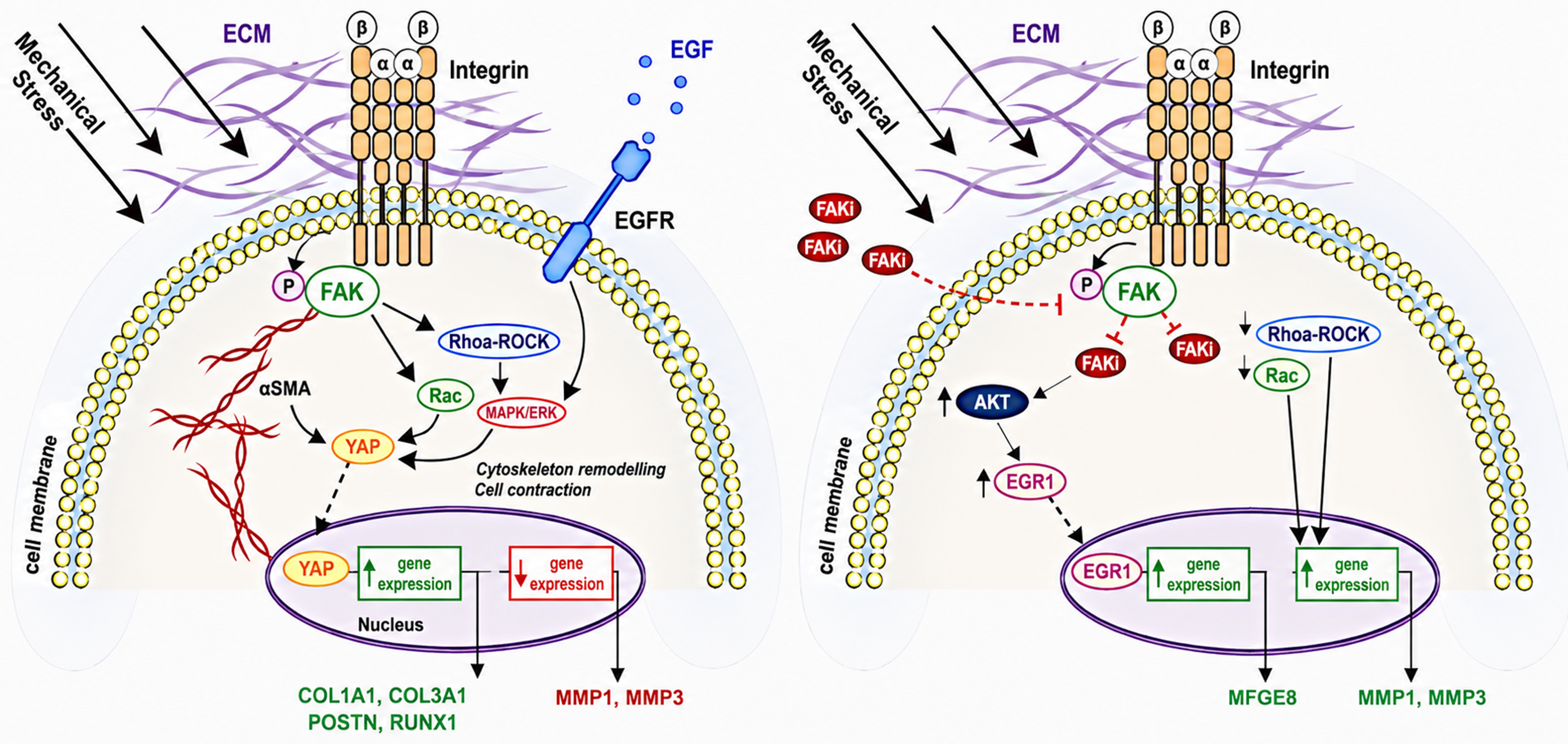
Figure 2. Schematic representation of FAK-dependent mechanotransduction and the effect of mechanical stress. (A) showing the activation of FAK-YAP signaling that promotes fibrosis (B) showing FAK inhibition and the activation of AKT-mediated induction of EGR1 and MFGE8, promoting regenerative repair. Adapted from Chen et al., Nature Communications, 2021[59], licensed under CC BY 4.0. ↑: Increased/upregulated; ↓: decreased/downregulated. ECM: Extracellular matrix; EGF: epidermal growth factor; EGFR: epidermal growth factor receptor; FAK: focal adhesion kinase; FAKi: focal adhesion kinase inhibitor; αSMA: alpha-smooth muscle actin; YAP: Yes-associated protein; AKT: protein kinase B; EGR1: early growth response 1; MAPK: mitogen-activated protein kinase; ROCK: Rho-associated coiled-coil containing protein kinase; Rac: Ras-related C3 botulinum toxin substrate; COL1A1: collagen type I alpha 1 chain; COL3A1: collagen type III alpha 1 chain; POSTN: periostin; RUNX1: runt-related transcription factor 1; MFGE8: milk fat globule-EGF factor 8; MMP1: matrix metalloproteininase 1; MMP3: matrix metalloproteininase 3.
Deletion of FAK in fibroblasts led to a marked reduction in inflammation, fibrosis, and myofibroblast activation, demonstrating the fibroblast-specific requirement for FAK in scar progression. Inhibition of FAK also activated an AKT-mediated regenerative pathway [Figure 2]. Furthermore, pharmacological inhibition of FAK in both in vitro human fibroblast cultures and in vivo animal models significantly suppressed MCP-1 secretion, inflammatory cell infiltration, ECM deposition, and subsequent scar formation. Collectively, these findings underscore the central role of the FAK-ERK-MCP-1 axis in mechanically induced fibrosis and suggest that targeting FAK may offer a promising therapeutic avenue to mitigate pathologic scarring and promote regenerative healing[54].
Additionally, targeting FAK may improve scar formation by other cellular processes as well, such as through their interaction with myeloid cells. A recent study by co-authors Chen et al., based on animal models, found that beyond the structural cells, immune cells are also mechanosensitive to external stimuli[60]. This research demonstrated that a subset of monocytes/macrophages sense the mechanical state of the environment and can drive fibrosis in the tissue. The mechanotransduction pathways via FAK in these immune cells are shown to regulate their differentiation, and targeting these pathways or recruiting these immune cells may reduce fibrosis[60]. Chen and colleagues explored how myeloid cells sense and respond to mechanical cues to regulate wound healing. The distinct mechanoresponsive myeloid subpopulations in both scars and unwounded skin that contribute to fibrotic outcomes were first identified. After applying exogenous mechanical strain in vivo and in vitro, it was found that selectively inhibiting myeloid mechanosignaling via the FAK pathway reduced the profibrotic myeloid subsets to promote native, anti-inflammatory populations. In wild-type mice, mechanically strained wounds produced pronounced scar formation with collagen fibers that were longer and highly aligned compared to unstrained controls. In contrast, myeloid-specific FAK knockout mice exhibited significantly narrower scars with shorter, more randomly organized collagen fibers resembling the natural basket-weave architecture of healthy skin[60].
Targeting FAK and the associated signaling pathways have also been seen with prior use of FAK inhibitors (FAKI), which have shown some promising results[61]. In an animal study by Ma et al., FAKI were incorporated into a dermal-like pullulan-collagen hydrogel matrix and showed optimistic results with murine excisional and burn wound models[62]. Small molecule FAKIs, such as VS-6062, were used originally in solid tumors but discontinued as antitumor agents due to systemic side effects when administered orally. However, intradermal administration of this molecule in small wounds as well as topical administration in larger wounds have shown benefits of reducing scar formation. Additionally, rapid and sustained release mechanisms of these molecules topically have shown promising use with burns and wounds that require more frequent dressing changes as well as splinted wounds[62]. FAKI hydrogels were also studied in preclinical settings while utilizing split thickness skin grafts (STSG) following burns. STSGs are at risk of HTS and contracture with enhanced fibroproliferative and inflammatory factors and upregulated mechanotransduction pathways[63]. In this study, FAKI was shown to reduce scar contraction and improve the biomechanical properties in STSG animal models compared to non-treated wounds[63]. Ultimately, FAKIs have shown overall faster wound repair and attenuation of scar formation, while also producing structurally stronger regenerated skin[62].
Other pathways of interest
Another seemingly important pathway for fibrogenesis and scar formation that is affected by mechanical forces is TGF-β/Smad. Transforming growth factor-β1 (TGF-β1) is a cytokine ligand that is expressed in states of inflammation by damaged tissue, causing receptor activation and downstream release of Smad transcriptional complexes that translocate and stimulate gene transcription[64-66]. As a result, fibroblasts are directly activated to produce collagen as well as induce differentiation into myofibroblasts[67,68]. Mechanical stress seems to have a role in this process: increased tensile force on myofibroblasts and ECM can release additional TGF-β1 and contribute to a positive feedback loop that results in excessive scar tissue formation[69]. TGF-β has also been shown to inhibit fibroblast/myofibroblast apoptosis as well as increase proliferation, resulting in sustained production of ECM proteins and superabundant scarring[67,70].
Another canonical molecular pathway for scar tissue formation that is potentiated by mechanical stress involves PDGF, a signaling protein that plays a significant role in normal wound healing. PDGF is directly involved with initiation of downstream signaling cascades that result in fibroblast chemotaxis and proliferation, stimulation of ECM protein production, and differentiation into myofibroblasts[71-74]. A number of additional pathways are actuated by PDGF signaling. The PI3K/AKT pathway drives cell survival and collagen/fibronectin synthesis via mTOR-dependent translation[75-78]. The MAPK pathway functions via the RAS/RAF/MEK/ERK cascade, a signaling module that serves to regulate fibroblast proliferation, myofibroblast differentiation, and ECM gene expression[79-81]. There is also significant cross-talk between these and other related pathways, leading to a complex environment with multiple levels of input. While PDGF expression normally peaks early in the wound-healing process, sustained production is a hallmark of dysregulated scarring[19,74,82,83].
Another notable player in the signaling cascade downstream from FAK is the YAP. When mechanical stress is detected by mechanosensitive ion channels, this leads to calcium influx and subsequent nuclear translocation of YAP [Figure 2]. This YAP activation has been shown to promote the transcription of profibrotic genes and stimulate ECM production, as demonstrated by hypertrophic scars in animal models. Inhibition of YAP, either pharmacologically or genetically, reduces scar formation and shifts wound healing toward a regenerative phenotype. This was seen in a co-authored study by Mascharak et al., which showed that by inhibiting the YAP mechanotransduction pathway in large animal models, fibroblast dynamics were altered[84]. The single use of Verteporfin, a pharmacological YAP inhibitor, following an injury was shown to reduce scar formation and restore tissue architecture[84]. Additional YAP inhibition was achieved through transgenic YAP knockout. Focusing on fibroblast subpopulations such as Engrailed-1 lineage positive and negative cells, inhibition of the YAP pathway as a means to block mechanotransduction signaling led to promotion of a more regenerative wound healing response that resulted in decreased fibrogenesis and scarring[85]. Further work on this topic elucidated the role of YAP signaling and mechanotransduction in the scarring process: by studying transcriptional and tissue structural changes resulting from YAP inhibition, additional signaling pathways (Trps1 and Wnt) were identified as key to the promotion of regenerative trajectories of wound healing rather than scar tissue formation[86].
Another pathway is the Wnt/β-catenin pathway in cutaneous scar formation. The Wnt pathway has been extensively studied over the past two decades, and it has emerged as another key element in the response of wound healing and scar formation to mechanical stimuli. The Wnt pathway has been shown to induce dermal fibrosis by affecting the cellular pathways leading to fibrosis, especially fibroblasts[87-89]. The enhanced β-catenin has been shown to increase collagen I gene expression and proliferation which in turn leads to scar formation[90,91]. It is also worth noting that fibroblasts expressing increased amounts of β-catenin have been found in hypertrophic scars and keloids[92]. The mechanical forces and stress play a role in increasing the transcription and translocation of the β-catenin in the nucleus[93,94]. Blocking of the activation of the Wnt pathway in vivo studies has shown promising results in decreasing skin fibrosis[89].
All of these cellular and molecular pathways affected by mechanical forces represent potential targets for pharmacological modulation in order to influence scar formation. Significant effort has gone into developing and commercializing various products to positively influence the scarring process. The economic burden of patients seeking scar revision far exceeds the cost of commercial products aimed at preventing or treating scars. This disparity has driven this significant interest in developing more effective strategies to modulate scar formation.
PRODUCT UTILIZATION OF MECHANOTRANSDUCTION IN SCAR FORMATION
Wong et al. studied the use of a stress shielding device around incisional scars[61]. The hypothesis was that forming an elastomeric polymer shielding device would prevent excessive scar formation by altering the mechanical environment of the wound. Red Duroc pigs were studied, which were considered ideal models to mimic human hypertrophic scars, as well as early clinical human models to study the safety and efficacy of these stress-shielding polymer devices. Excisional full-thickness wounds were created on red Duroc pigs and followed for 8 weeks with the polymer device and were compared with the unshielded incisions. The results showed decreased scar hypertrophy when using the shielding device, with characteristics similar to unwounded skin. The device was then used on human patients following abdominoplasty and showed similar results at 6-12 month follow-up with improved scar appearance judged by an expert panel[61]. FAK inhibition has also demonstrated measurable improvements in the biomechanical properties of healed skin. In preclinical wound models, FAKI treated wounds showed increased Young’s modulus and ultimate measured tensile strength compared to controls, indicating improved tissue strength, structural integrity, and resistance to mechanical stress[62]. YAP inhibition with verteporfin in large animal models demonstrated measurable improvements in wound biomechanics. Verteporfin-treated wounds regained elasticity over time and exhibited ultimate tensile strength and Young’s modulus comparable to unwounded skin by 16 weeks, whereas control wounds remained stiffer and mechanically weaker, suggesting a more regenerative healing phenotype[84].
Other products aimed at decreasing tension around wounds to reduce scar formation include surgical tapes[95], force modulating tissue bridges (FMTB)[96], and silicone gel sheets[97], as well as other surgical closure techniques utilizing sutures to offload mechanical tension. Silicone gel sheets are one of the most widely used adjuncts to reduce scars in hypertrophic scars and keloids since the 1980s. One way the silicone gel sheet works is by hydration and occlusion of the incision. However, it is also thought that the silicone distributes the forces from the wound borders into the sheet itself due to its elasticity. This reduces the mechanical tension, thereby reducing collagen deposition and scar formation[95]. Additionally, targeting the subset of immune mechanosensing monocytes/macrophages may open the possibility of interventions before ECM fibrosis is established by modulating immune mechanosensing rather than only targeting fibroblasts/collagen[60] [Table 1].
Bench-to-bedside table that maps each mechanomodulatory strategy to (a) wound-healing phase targeted; (b) hypothesized mechanism; and (c) level of evidence and outcomes
| Mechanomodulatory strategy | Wound healing phase targeted | Hypothesized mechanism of action | Level of evidence and outcomes | Key Ref. |
| Stress shielding (silicon gel, surgical tapes) | Proliferative | Reduced mechanical tension, thereby reducing collagen deposition and myofibroblast formation | Preclinical animal studies and human clinical trials, reduced pigmentation and height of scars formed | [61,95] |
| Closure techniques (Z-plasty, deep dermal, layered closure) | Proliferative | Redistributes tension across the incision, thereby reducing profibrotic pathways | Human trials, improved overall cosmesis of scars | [45,47,48] |
| FAK inhibition | Inflammatory + Proliferative | Modulates immune response to mechanotransduction and shifts macrophage phenotype away from profibrotic. Also suppresses profibrotic pathways | Preclinical animal studies, reduced profibrotic myeloid population | [58-60,62,63] |
| YAP/TAZ inhibition | Proliferative | Blocks fibroblast proliferation | Preclinical animal studies, reduced scar thickness | [84-86] |
| Wnt/β-catenin inhibition | Proliferative | Inhibits α-SMA expression | Preclinical, reduces myofibroblast differentiation | [89] |
There still appears to be great interest in reducing scars in the plastic surgery field, and understanding the mechanism of these scars continues to develop. These products continue to be actively investigated as they underscore the importance of mechanotransduction in reducing scar formation, particularly in hypertrophic scars and keloids. Importantly, as we continue to understand the cellular pathophysiology of scar formation, these strategies will emphasize the prevention rather than revision of scars, which shifts scar management toward early intervention in high-risk wounds.
LIMITATIONS AND CONTROVERSIES
Although substantial research has investigated the effects of mechanotransduction on hypertrophic scar formation, a number of limitations exist. As evidenced by the literature cited by this paper, the majority of the studied pathways and effects of mechanical tension on scarring were investigated using animal models, particularly murine. While we have made an effort to incorporate particular animal models that better mimic human scarring, such as red Duroc pigs, a lack of adequately powered human trials remains a significant limitation within this field. The nature of modern scientific research necessitates the sequential progression from in vitro to in vivo and finally human trials, meaning the available research will be carried out using mostly cell and animal models. This is reflected in the studies utilized in writing this paper. Ethics, ease of access, and availability of reagents all contribute to greater availability of animal and cell studies. Human trials remain the gold standard of research, and it is important to keep in mind that the translational potential of animal studies will always be somewhat limited due to interspecies differences. Further human trials may reveal additional molecular idiosyncrasies that are unique and inherently involved with human scar formation.
Conflicting research results also serve to illustrate the missing links in our understanding of the molecular basis of human scarring. While many studies demonstrate that increased tensile forces promote scar formation via activation of profibrotic pathways, other published investigations suggest that mechanotransduction responses are highly context-dependent and vary according to wound timing, ECM stiffness, fibroblast lineage heterogeneity, and the balance between regenerative and fibrotic signaling pathways. These inconsistencies likely reflect a dynamic wound-healing process that is regulated by a combination of chemical, mechanical, and other forces.
Finally, many of the products studied and applied to human subjects have so far shown mixed results. There is no single product that provides definitive reduction in human scar formation. Additional randomized human clinical trials will be required to study these products and establish the clear causation and consistent results to continue with further application of such products.
CONCLUSION
Scar formation remains an inevitable outcome of wound healing, especially in high tension areas of the body. It is increasingly evident that mechanical forces play a pivotal role in dictating the quality and extent of this complex process. From the alignment of Langer’s lines to the cellular signaling cascades of FAK, TGF-β/Smad, PDGF, YAP, and Wnt/β-catenin, mechanical cues have been shown to directly shape fibroblast behavior, collagen deposition, and ultimately the architecture of scar tissue. A deeper understanding of mechanotransduction has not only clarified the pathophysiology of hypertrophic and keloid scars but has also opened new therapeutic avenues. Clinical strategies that respect skin biomechanics, such as incision planning, stress-shielding devices, and novel pharmacologic inhibitors, are now showing promise in reducing pathological scarring and improving functional and cosmetic outcomes. Although scarring is often perceived as a primarily cosmetic issue, the reality is that many patients are affected both mentally and physically by what they deem to be unsightly features or imperfections, especially when dealing with pathologic processes such as HTS or keloids. The cost of treatment is high, ranging from several hundred dollars for topical therapies to several thousand for surgical scar revision[98-100]. Discovering methods to prevent these problems before they arise is both meaningful to patients and economically worthwhile for the healthcare system. Future research should prioritize clinical studies investigating mechanotransduction-targeted therapies and tension-modulating strategies in wound healing. Advancements in regenerative medicine and mechanobiology may further support the development of personalized approaches to reduce fibrosis and improve scar outcomes.
DECLARATIONS
Authors’ contributions
Made substantial contributions to the conception of the review and drafted the initial manuscript and contributed to literature acquisition and critical revision of the manuscript: Sheckley M, Lowney M, Hahn WW
Contributed to the conceptual framework of the review, provided expert interpretation of mechanotransduction pathways, and critically revised the manuscript for important intellectual content: Chen K, Gurtner GC
All authors read and approved the final manuscript.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool ChatGPT (OpenAI, GPT-5.5, released 2026-04-24) was used solely for limited editorial assistance, including phrasing refinement, grammar correction, paraphrasing, and improvement of sentence clarity and readability. In addition, The Graphic Abstract was also created with the assistance of ChatGPT (OpenAI, GPT-5.5, released 2026-04-24) with similar phrasing and set up with the figures created using tools of Microsoft PowerPoint (Version 16.109.3). The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
None.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
4. Willenborg S, Injarabian L, Eming SA. Role of macrophages in wound healing. Cold Spring Harb Perspect Biol. 2022;14:a041216.
6. Trace AP, Enos CW, Mantel A, Harvey VM. Keloids and hypertrophic scars: a spectrum of clinical challenges. Am J Clin Dermatol. 2016;17:201-23.
7. Fang X, Wang Y, Chen H, et al. Hypertrophic scarring and keloids: epidemiology, molecular pathogenesis, and therapeutic interventions. MedComm. 2025;6:e70381.
8. Anderson JB, Foglio A, Harrant AB, et al. Scoping review of therapeutic strategies for keloids and hypertrophic scars. Plast Reconstr Surg Glob Open. 2021;9:e3469.
9. De Faverney PM, Molamodi K, Tancrede-Bohin E, Verschoore M. Support for dermatological research in Sub-Saharan Africa: insights from African hair and skin research programs. Int J Dermatol. 2024;63:1081-8.
10. Nangole FW, Agak GW. Keloid pathophysiology: fibroblast or inflammatory disorders? JPRAS Open. 2019;22:44-54.
11. Berman B, Maderal A, Raphael B. Keloids and hypertrophic scars: pathophysiology, classification, and treatment. Dermatol Surg. 2017;43:S3-S18.
12. Chike-Obi CJ, Cole PD, Brissett AE. Keloids: pathogenesis, clinical features, and management. Semin Plast Surg. 2009;23:178-84.
13. Brissett AE, Sherris DA. Scar contractures, hypertrophic scars, and keloids. Facial Plast Surg. 2001;17:263-72.
14. Zhou B, Gao Z, Liu W, Wu X, Wang W. Important role of mechanical microenvironment on macrophage dysfunction during keloid pathogenesis. Exp Dermatol. 2022;31:375-80.
15. Tsai CH, Ogawa R. Keloid research: current status and future directions. Scars Burn Heal. 2019;5:2059513119868659.
16. Ogawa R, Okai K, Tokumura F, et al. The relationship between skin stretching/contraction and pathologic scarring: the important role of mechanical forces in keloid generation. Wound Repair Regen. 2012;20:149-57.
17. Liu S, Yang H, Song J, Zhang Y, Abualhssain ATH, Yang B. Keloid: genetic susceptibility and contributions of genetics and epigenetics to its pathogenesis. Exp Dermatol. 2022;31:1665-75.
18. Kim HJ, Kim YH. Comprehensive insights into keloid pathogenesis and advanced therapeutic strategies. Int J Mol Sci. 2024;25:8776.
19. Mony MP, Harmon KA, Hess R, Dorafshar AH, Shafikhani SH. An updated review of hypertrophic scarring. Cells. 2023;12:678.
20. Limandjaja GC, Niessen FB, Scheper RJ, Gibbs S. Hypertrophic scars and keloids: overview of the evidence and practical guide for differentiating between these abnormal scars. Exp Dermatol. 2021;30:146-61.
21. Choi C, Mukovozov I, Jazdarehee A, et al. Management of hypertrophic scars in adults: a systematic review and meta-analysis. Australas J Dermatol. 2022;63:172-89.
22. Bailey J, Schwehr M, Beattie A. Management of keloids and hypertrophic scars. Am Fam Physician. 2024;110:605-11.
23. Elsaie ML. Update on management of keloid and hypertrophic scars: a systemic review. J Cosmet Dermatol. 2021;20:2729-38.
24. Peña OA, Martin P. Cellular and molecular mechanisms of skin wound healing. Nat Rev Mol Cell Biol. 2024;25:599-616.
26. Sorg H, Tilkorn DJ, Hager S, Hauser J, Mirastschijski U. Skin wound healing: an update on the current knowledge and concepts. Eur Surg Res. 2017;58:81-94.
27. Mouw JK, Ou G, Weaver VM. Extracellular matrix assembly: a multiscale deconstruction. Nat Rev Mol Cell Biol. 2014;15:771-85.
28. Rousselle P, Montmasson M, Garnier C. Extracellular matrix contribution to skin wound re-epithelialization. Matrix Biol. 2019;75-76:12-26.
29. Pakyari M, Farrokhi A, Maharlooei MK, Ghahary A. Critical role of transforming growth factor beta in different phases of wound healing. Adv Wound Care. 2013;2:215-24.
30. Roberts AB, Flanders KC, Heine UI, et al. Transforming growth factor-beta: multifunctional regulator of differentiation and development. Philos Trans R Soc Lond B Biol Sci. 1990;327:145-54.
31. Juhl P, Bondesen S, Hawkins CL, et al. Dermal fibroblasts have different extracellular matrix profiles induced by TGF-β, PDGF and IL-6 in a model for skin fibrosis. Sci Rep. 2020;10:17300.
32. Fang X, Hu X, Zheng Z, et al. Smad interacting protein 1 influences transforming growth factor-β1/Smad signaling in extracellular matrix protein production and hypertrophic scar formation. J Mol Histol. 2019;50:503-14.
33. Desmoulière A, Chaponnier C, Gabbiani G. Tissue repair, contraction, and the myofibroblast. Wound Repair Regen. 2005;13:7-12.
34. Darby IA, Laverdet B, Bonté F, Desmoulière A. Fibroblasts and myofibroblasts in wound healing. Clin Cosmet Investig Dermatol. 2014;7:301-11.
35. Ibrahim MM, Chen L, Bond JE, et al. Myofibroblasts contribute to but are not necessary for wound contraction. Lab Invest. 2015;95:1429-38.
36. Barnes LA, Marshall CD, Leavitt T, et al. Mechanical forces in cutaneous wound healing: emerging therapies to minimize scar formation. Adv Wound Care. 2018;7:47-56.
37. Junker JP, Kratz C, Tollbäck A, Kratz G. Mechanical tension stimulates the transdifferentiation of fibroblasts into myofibroblasts in human burn scars. Burns. 2008;34:942-6.
38. Hosgood G. Stages of wound healing and their clinical relevance. Vet Clin North Am Small Anim Pract. 2006;36:667-85.
39. Rognoni E, Pisco AO, Hiratsuka T, et al. Fibroblast state switching orchestrates dermal maturation and wound healing. Mol Syst Biol. 2018;14:e8174.
40. Marshall CD, Hu MS, Leavitt T, Barnes LA, Lorenz HP, Longaker MT. Cutaneous scarring: basic science, current treatments, and future directions. Adv Wound Care. 2018;7:29-45.
41. Gurtner GC, Werner S, Barrandon Y, Longaker MT. Wound repair and regeneration. Nature. 2008;453:314-21.
42. Frech FS, Hernandez L, Urbonas R, Zaken GA, Dreyfuss I, Nouri K. Hypertrophic scars and keloids: advances in treatment and review of established therapies. Am J Clin Dermatol. 2023;24:225-45.
43. Betarbet U, Blalock TW. Keloids: a review of etiology, prevention, and treatment. J Clin aesthetic Dermatol. 2020;13:33-43.
44. Huang C, Quong WL, Kamii Y, Ogawa R. Ideal surgical incision lines minimizing tension: a proposal based on observations of hypertrophic scars and keloids. Plast Reconstr Surg Glob Open. 2025;13:e7344.
45. Wray RC. Force required for wound closure and scar appearance. Plast Reconstr Surg. 1983;72:380-2.
46. Gurtner GC, Dauskardt RH, Wong VW, et al. Improving cutaneous scar formation by controlling the mechanical environment: large animal and phase I studies. Ann Surg. 2011;254:217-25.
49. Berry CE, Downer M Jr, Morgan AG, et al. The effects of mechanical force on fibroblast behavior in cutaneous injury. Front Surg. 2023;10:1167067.
50. Halper J. Basic components of connective tissues and extracellular matrix: fibronectin, fibrinogen, laminin, elastin, fibrillins, fibulins, matrilins, tenascins and thrombospondins. Adv Exp Med Biol. 2021;1348:105-26.
51. Ma Y, Feng X, Rogers JA, Huang Y, Zhang Y. Design and application of ‘J-shaped’ stress-strain behavior in stretchable electronics: a review. Lab Chip. 2017;17:1689-704.
52. Ling S, Zhang Q, Kaplan DL, Omenetto F, Buehler MJ, Qin Z. Printing of stretchable silk membranes for strain measurements. Lab Chip. 2016;16:2459-66.
53. Fernandes MG, da Silva LP, Cerqueira MT, et al. Mechanomodulatory biomaterials prospects in scar prevention and treatment. Acta Biomater. 2022;150:22-33.
54. Kness-Knezinskis E, Sheckley M, Hostler AC, Mora Pinos MG, Chen K, Gurtner GC. Translational approaches manipulating mechanobiology to promote scarless healing in humans. J Plast Reconstr Aesthet Surg. 2026;112:25-33.
56. Wu C. Focal adhesion: a focal point in current cell biology and molecular medicine. Cell Adh Migr. 2007;1:13-8.
57. Tan X, Yan Y, Song B, Zhu S, Mei Q, Wu K. Focal adhesion kinase: from biological functions to therapeutic strategies. Exp Hematol Oncol. 2023;12:83.
58. Wong VW, Rustad KC, Akaishi S, et al. Focal adhesion kinase links mechanical force to skin fibrosis via inflammatory signaling. Nat Med. 2011;18:148-52.
59. Chen K, Kwon SH, Henn D, et al. Disrupting biological sensors of force promotes tissue regeneration in large organisms. Nat Commun. 2021;12:5256.
60. Chen K, Griffin M, Henn D, et al. Targeting circulating mechanoresponsive monocytes and macrophages to reduce fibrosis. Nat Biomed Eng. 2026;10:1247-62.
61. Wong VW, Beasley B, Zepeda J, et al. A mechanomodulatory device to minimize incisional scar formation. Adv Wound Care. 2013;2:185-94.
62. Ma K, Kwon SH, Padmanabhan J, et al. Controlled delivery of a focal adhesion kinase inhibitor results in accelerated wound closure with decreased scar formation. J Invest Dermatol. 2018;138:2452-60.
63. Chen K, Henn D, Januszyk M, et al. Disrupting mechanotransduction decreases fibrosis and contracture in split-thickness skin grafting. Sci Transl Med. 2022;14:eabj9152.
64. Kohlhauser M, Mayrhofer M, Kamolz LP, Smolle C. An update on molecular mechanisms of scarring-a narrative review. Int J Mol Sci. 2024;25:11579.
65. Hu HH, Chen DQ, Wang YN, et al. New insights into TGF-β/Smad signaling in tissue fibrosis. Chem Biol Interact. 2018;292:76-83.
66. Zhang T, Wang XF, Wang ZC, et al. Current potential therapeutic strategies targeting the TGF-β/Smad signaling pathway to attenuate keloid and hypertrophic scar formation. Biomed Pharmacother. 2020;129:110287.
67. Shi X, Young CD, Zhou H, Wang X. Transforming growth factor-β signaling in fibrotic diseases and cancer-associated fibroblasts. Biomolecules. 2020;10:1666.
68. Dobaczewski M, Bujak M, Li N, et al. Smad3 signaling critically regulates fibroblast phenotype and function in healing myocardial infarction. Circ Res. 2010;107:418-28.
69. Wipff PJ, Rifkin DB, Meister JJ, Hinz B. Myofibroblast contraction activates latent TGF-beta1 from the extracellular matrix. J Cell Biol. 2007;179:1311-23.
70. Bao H, Jiang K, Meng K, et al. TGF-β2 induces proliferation and inhibits apoptosis of human Tenon capsule fibroblast by miR-26 and its targeting of CTGF. Biomed Pharmacother. 2018;104:558-65.
71. Jung SC, Kang D, Ko EA. Roles of PDGF/PDGFR signaling in various organs. Korean J Physiol Pharmacol. 2025;29:139-55.
73. Irma J, Kartasasmita AS, Kartiwa A, Irfani I, Rizki SA, Onasis S. From growth factors to structure: PDGF and TGF-β in granulation tissue formation. A literature review. J Cell Mol Med. 2025;29:e70374.
74. Rajkumar VS, Shiwen X, Bostrom M, et al. Platelet-derived growth factor-beta receptor activation is essential for fibroblast and pericyte recruitment during cutaneous wound healing. Am J Pathol. 2006;169:2254-65.
75. He X, Li Y, Deng B, et al. The PI3K/AKT signalling pathway in inflammation, cell death and glial scar formation after traumatic spinal cord injury: mechanisms and therapeutic opportunities. Cell Prolif. 2022;55:e13275.
76. Zhao S, Liu H, Wang H, et al. Inhibition of phosphatidylinositol 3-kinase catalytic subunit alpha by miR-203a-3p reduces hypertrophic scar formation via phosphatidylinositol 3-kinase/AKT/mTOR signaling pathway. Burns Trauma. 2024;12:tkad048.
77. Jere SW, Houreld NN, Abrahamse H. Role of the PI3K/AKT (mTOR and GSK3β) signalling pathway and photobiomodulation in diabetic wound healing. Cytokine Growth Factor Rev. 2019;50:52-9.
78. Teng Y, Fan Y, Ma J, et al. The PI3K/Akt pathway: emerging roles in skin homeostasis and a group of non-malignant skin disorders. Cells. 2021;10:1219.
79. Darby IA, Desmoulière A. Scar formation: cellular mechanisms. In: Téot L, Mustoe TA, Middelkoop E, Gauglitz GG, Editors. Textbook on scar management: state of the art management and emerging technologies. Cham (CH): Springer; 2020.
80. Bahar ME, Kim HJ, Kim DR. Targeting the RAS/RAF/MAPK pathway for cancer therapy: from mechanism to clinical studies. Signal Transduct Target Ther. 2023;8:455.
81. Umbarkar P, Tousif S, Singh AP, et al. Fibroblast GSK-3α promotes fibrosis via RAF-MEK-ERK pathway in the injured heart. Circ Res. 2022;131:620-36.
82. Pierce GF, Mustoe TA, Altrock BW, Deuel TF, Thomason A. Role of platelet‐derived growth factor in wound healing. J of Cellular Biochemistry. 1991;45:319-26.
84. Mascharak S, Griffin M, Talbott HE, et al. Inhibiting mechanotransduction prevents scarring and yields regeneration in a large animal model. Sci Transl Med. 2025;17:eadt6387.
85. Mascharak S, desJardins-Park HE, Davitt MF, et al. Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring. Science. 2021;372:eaba2374.
86. Mascharak S, Talbott HE, Januszyk M, et al. Multi-omic analysis reveals divergent molecular events in scarring and regenerative wound healing. Cell Stem Cell. 2022;29:315-27.e6.
87. Jin X, Wang J, Cao R, Jiang D. Wnt signaling pathway: biological function, diseases, and therapeutic interventions. MedComm. 2026;7:e70580.
88. Mullin NK, Mallipeddi NV, Hamburg-Shields E, Ibarra B, Khalil AM, Atit RP. Wnt/β-catenin signaling pathway regulates specific lncRNAs that impact dermal fibroblasts and skin fibrosis. Front Genet. 2017;8:183.
89. Griffin MF, Huber J, Evan FJ, Quarto N, Longaker MT. The role of Wnt signaling in skin fibrosis. Med Res Rev. 2022;42:615-28.
90. Bielefeld KA, Amini-Nik S, Alman BA. Cutaneous wound healing: recruiting developmental pathways for regeneration. Cell Mol Life Sci. 2013;70:2059-81.
91. Hamburg-Shields E, DiNuoscio GJ, Mullin NK, Lafyatis R, Atit RP. Sustained β-catenin activity in dermal fibroblasts promotes fibrosis by up-regulating expression of extracellular matrix protein-coding genes. J Pathol. 2015;235:686-97.
92. Sato M. Upregulation of the Wnt/beta-catenin pathway induced by transforming growth factor-beta in hypertrophic scars and keloids. Acta Derm Venereol. 2006;86:300-7.
93. Premaraj S, Souza I, Premaraj T. Mechanical loading activates β-catenin signaling in periodontal ligament cells. Angle Orthod. 2011;81:592-9.
94. Mendez MG, Janmey PA. Transcription factor regulation by mechanical stress. Int J Biochem Cell Biol. 2012;44:728-32.
95. O’Reilly S, Crofton E, Brown J, Strong J, Ziviani J. Use of tape for the management of hypertrophic scar development: a comprehensive review. Scars Burn Heal. 2021;7:20595131211029206.
96. Kazmer DO, Eaves FF 3rd. Force modulating tissue bridges for reduction of tension and scar: finite element and image analysis of preclinical incisional and nonincisional models. Aesthet Surg J. 2018;38:1250-63.
97. Bleasdale B, Finnegan S, Murray K, Kelly S, Percival SL. The use of silicone adhesives for scar reduction. Adv Wound Care. 2015;4:422-30.
98. Block L, Gosain A, King TW. Emerging therapies for scar prevention. Adv Wound Care. 2015;4:607-14.
99. McPhail SM, Wiseman J, Simons M, Kimble R, Tyack Z. Cost-effectiveness of scar management post-burn: a trial-based economic evaluation of three intervention models. Sci Rep. 2022;12:18601.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].