


Imidazole propionate: a new direction for atherosclerosis prevention and treatment
 0
0 From lipid accumulation to gut-derived immunometabolic regulation
Atherosclerotic cardiovascular disease (ASCVD) has long been considered a lipid-driven disease, with low-density lipoprotein cholesterol (LDL-C) representing its central modifiable risk factor[1]. Despite the success of lipid-lowering therapies, substantial residual cardiovascular risk remains, indicating that additional pathogenic mechanisms contribute to disease progression[1]. Recently, immune dysregulation and metabolic inflammation have emerged as independent, clinically relevant drivers of atherosclerosis[2]. Metabolites produced by gut microbiota are now recognised for their potential to influence vascular pathology by modulating immune and metabolic processes. In a recent article published in Nature, Mastrangelo et al. identify microbially produced imidazole propionate (ImP), previously linked to metabolic disorders, as a cholesterol-independent driver of atherosclerosis[3].
Causal evidence linking ImP to atherosclerosis
Existing studies suggest that ImP is associated with heart failure and mortality, potentially mediated through chronic systemic inflammation and immune activation[4]. Nonetheless, its predictive value for atherosclerotic progression and its direct role in the development of atherosclerotic disease have not been fully established. Mastrangelo et al. conducted diet-based screening in Apoe-/- and Ldlr-/- mice, identifying ImP and demonstrating its association with aortic arch lesion under high-cholesterol conditions[3]. This association was subsequently extended to human cohorts, in which circulating ImP levels were linked to the presence, severity, and activity of subclinical atherosclerosis. Notably, ImP concentrations exhibited a clear dose-response relationship with subclinical atherosclerotic endpoints and showed additive value when combined with established blood-derived atherosclerosis biomarkers[3].
Causality is further strengthened by a series of interventional experiments at both the animal and cellular levels. Exogenous ImP supplementation accelerates atherosclerosis progression in Apoe-/- mice without affecting circulating cholesterol or glucose levels, thereby excluding major metabolic confounders and supporting a direct pathogenic role. Importantly, these effects were shown to be reversible: pharmacological inhibition of the ImP-binding imidazoline receptor 1 (I1R) with AGN192403 markedly attenuates atherosclerotic lesion development[3].
Immunometabolic links ImP to vascular inflammation
Beyond establishing causality, the study offers mechanistic insight into how ImP promotes a pro-atherogenic inflammatory milieu through immunometabolic regulation. Single-cell transcriptomic analyses link ImP exposure to enhanced systemic and vascular inflammation[3]. After entering the circulation, ImP preferentially targets myeloid cells and engages the I1R at the cell surface. Activation of I1R triggers downstream mTOR-centred signalling, reflected by increased phosphorylation of S6 (p-S6), and drives a sustained pro-inflammatory immune programme. This immunometabolic shift is accompanied by increased production of cytokines such as TNF and IFN-γ, expansion of circulating Ly6Chi monocytes, enhanced T-cell activation, and accumulation of inflammatory macrophages within the vascular wall [Figure 1]. Significantly, both genetic disruption and pharmacological inhibition of I1R abrogate ImP-induced p-S6 activation and inflammatory cytokine production, thereby placing I1R upstream of these immunometabolic effects[3]. Thus, ImP may serve as a molecular link between gut-derived metabolism, immune activation, vascular inflammation, and pathology in atherosclerosis.
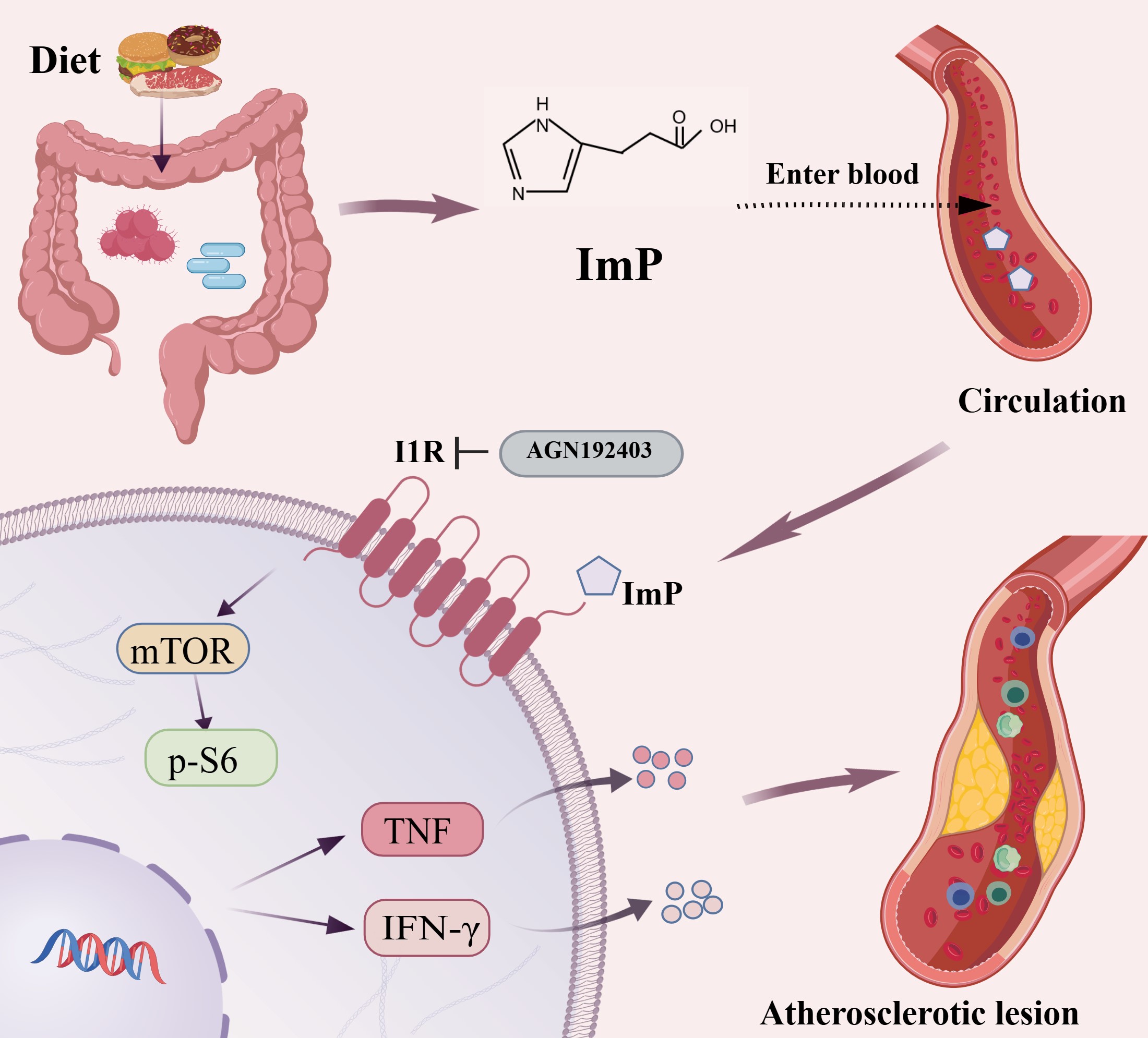
Figure 1. Gut-derived imidazole propionate (ImP) activates the imidazoline receptor 1 (I1R) and the mTOR pathway in myeloid cells, driving immune responses and vascular inflammation, which contribute to the progression of atherosclerosis. Created with BioGDP.com.
Animal models for evaluating gut-derived metabolic risk factors
While mouse models, such as Apoe-/- and Ldlr-/- strains, are widely used in atherosclerosis research due to their genetic tractability and mechanistic utility, they differ substantially from humans in gut anatomy, microbiota composition, and metabolic regulation. These differences limit their ability to model long-term, diet-dependent microbial metabolites. For example, Mastrangelo et al.'s study showed that the microbiota capable of producing ImP were different between humans and mice[3].
Rabbits show lipid metabolism and plaque features that more closely resemble those of humans and are useful for studying atherosclerotic lesion structure, although their herbivorous physiology restricts modelling of complex dietary effects on microbial metabolism[5]. In contrast, pigs are omnivorous and resemble humans in gastrointestinal structure, gut microbiota, lipid metabolism and coronary artery disease, making them well suited for preclinical studies of diet-microbiota-metabolite interactions. Non-human primates (NHPs) provide the closest approximation to humans in gastrointestinal physiology, immune regulation, metabolic homeostasis, and atherosclerotic lesions[6]. Therefore, they represent a valuable translational platform for validating the long-term vascular effects of metabolites in atherosclerosis. Thus, identifying animal models that better simulate human dietary diversity is critical for investigating diet-driven microbial metabolites in atherosclerosis.
Translational implications and future directions
ImP is strongly associated with early and active atherosclerosis and provides additional discriminatory value over LDL-C and high-sensitivity C-reactive protein alone, supporting its potential utility as a biomarker for early disease detection and refined risk assessment[3]. These findings also highlight opportunities for early prevention. Notably, inhibition of the ImP-I1R signalling axis attenuates atherosclerosis, and administration of the I1R antagonist AGN192403 rescues plaque progression under high-cholesterol dietary conditions[3], identifying a potential new therapeutic target for atherosclerosis. While AGN192403 serves as a useful experimental I1R antagonist, its utility remains confined to mechanistic studies, highlighting the need for clinically viable I1R-targeting agents. Additionally, I1R is widely expressed across multiple tissues and is involved in the regulation of behavioural processes and the pathogenesis of various diseases, including hypertension, obesity, and Huntington’s disease[7]. Consequently, the long-term safety of systemic I1R inhibition, as well as systemic effects, remains to be fully evaluated.
Against the backdrop of residual cardiovascular risk, ImP as a cholesterol-independent factor adds a new layer to our understanding of disease mechanisms[3]. ImP-mediated atherosclerosis further supports inflammation as an independent contributor to residual ASCVD risk. Notably, multiple studies have demonstrated that interventions targeting inflammatory pathways, such as low-dose colchicine and IL-1β inhibition, can reduce cardiovascular events in selected populations with ASCVD[2]. In this context, the ImP-I1R axis provides a new entry point for understanding inflammatory risk.
In considering the translational potential of the ImP-I1R axis, several important limitations warrant attention. First, gut microbial taxa linked to ImP only partially overlap between mice and humans, complicating cross-species extrapolation. Second, the association between ImP and cardiometabolic risk factors varies across human populations, indicating that its clinical relevance may be influenced by population-specific traits. Third, while human data reveal a significant sex-dependent difference in ImP levels, experimental studies have not systematically dissected sex-specific mechanisms. To address this gap, future studies should include balanced male and female mice with or without ImP supplementation. At the mechanistic level, further studies are warranted to determine whether estrogenic and androgenic signalling modulates the proatherogenic effects of ImP. Lastly, the study did not evaluate the relationship between ImP and other major gut-derived cardiovascular metabolites, particularly trimethylamine N-oxide (TMAO), leaving unclear whether ImP acts as an independent pathogenic driver or as part of a broader microbial-metabolic network. Notably, despite over a decade of robust validation in both preclinical and clinical studies, TMAO has not yet translated into approved therapies[8]. These translational challenges suggest that future evaluation of gut-derived metabolites in atherosclerosis should move beyond isolated associative findings and integrate cross-species reproducibility, population robustness, mechanistic specificity, and network-level interactions.
DECLARATIONS
Authors’ contributions
Conceptualised, drafted, and revised the manuscript: Ba Z, Liu E
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool ChatGPT (OpenAI, version based on GPT-5.2, released 2025-12-11) was used solely for language editing. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
This commentary was partially supported by the National Natural Science Foundation of China (No. 82270496).
Conflicts of interest
Both authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Hoogeveen RC, Ballantyne CM. Residual cardiovascular risk at low LDL: remnants, lipoprotein(a), and inflammation. Clin Chem. 2021;67:143-53.
2. Mensah GA, Arnold N, Prabhu SD, Ridker PM, Welty FK. Inflammation and cardiovascular disease: 2025 ACC scientific statement: a report of the American College of Cardiology. J Am Coll Cardiol. 2026;87:1381-404.
3. Mastrangelo A, Robles-Vera I, Mañanes D, et al. Imidazole propionate is a driver and therapeutic target in atherosclerosis. Nature. 2025;645:254-61.
4. Molinaro A, Nemet I, Bel Lassen P, et al. Microbially produced imidazole propionate is associated with heart failure and mortality. JACC Heart Fail. 2023;11:810-21.
5. Fan J, Kitajima S, Watanabe T, et al. Rabbit models for the study of human atherosclerosis: from pathophysiological mechanisms to translational medicine. Pharmacol Ther. 2015;146:104-19.
6. Jia L, Qu P, Zhao Y, et al. Tripeptide DT-109 (Gly-Gly-Leu) attenuates atherosclerosis and vascular calcification in nonhuman primates. Sig Transduct Target Ther. 2025;10:122.
7. Bousquet P, Hudson A, García-Sevilla JA, Li J. Imidazoline receptor system: the past, the present, and the future. Pharmacol Rev. 2020;72:50-79.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].