


Cardiovascular aging through the lens of Blue Zones
 0
0  ,
, Abstract
Blue Zones are geographically distinct populations characterized by exceptional longevity and remarkably low rates of cardiovascular disease. Although these regions differ culturally and environmentally, they share common lifestyle patterns, including habitual natural physical activity, predominantly plant-based diets, caloric moderation, strong social integration, stress reduction, and purpose-driven living. In this review, we examine the cardiovascular phenotype of Blue Zone populations and discuss the biologic mechanisms through which these lifestyle patterns may influence vascular aging, metabolic health, and longevity. We summarize emerging evidence linking Blue Zone-associated behaviors to improved endothelial function, reduced arterial stiffness, mitochondrial quality control, attenuation of chronic inflammation, and preservation of metabolic flexibility. Focus is placed on nutrient-sensing pathways involving adenosine monophosphate-activated protein kinase, Sirtuin 1, Forkhead box O signaling, and mTOR, as well as mechanisms related to autophagy, oxidative stress, inflammation, endothelial homeostasis, and cellular aging. We further review evidence regarding immune regulation, epigenetic aging, telomere stability, and longevity-associated biomarkers observed in centenarian populations. Finally, we discuss the translational implications of the Blue Zones framework for preventive cardiology and public health, including the development of culturally adaptable lifestyle interventions to extend health span and reduce the burden of cardiovascular and age-related disease.
Keywords
INTRODUCTION
The global population is undergoing a rapid demographic shift towards older age, driven largely by advances in healthcare, medical technology, and improved nutrition[1]. According to a 2019 report by the United Nations, the number of individuals aged 65 years and older is projected to more than double from 703 million in 2019 to over 1.5 billion by 2050[2]. Notably, the population aged 80 years and older is expected to grow even more rapidly, increasing nearly threefold from 143 to 426 million over the same period[2]. Understanding the biological and lifestyle factors that promote healthy aging is crucial, particularly among centenarians who often maintain longevity without chronic disease[3-5]. Identifying these determinants may offer insight into strategies that enhance both lifespan and health span.
Dan Buettner, a National Geographic Fellow, first introduced the concept of "Blue Zones,” a term describing regions around the world with unusually high concentrations of centenarians [Figure 1]. These include Okinawa (Japan), Sardinia (Italy), the Nicoya Peninsula (Costa Rica), Ikaria (Greece), and Loma Linda (United States)[2,3]. The striking longevity observed in these areas, compared to global averages, has sparked significant scientific interest in uncovering the lifestyle and environmental factors that contribute to sustained healthy longevity. Buettner and his team sought to identify common behavioral and social determinants, such as patterns of physical activity, dietary habits, psychosocial well-being, and community engagement that might explain the exceptional longevity in these populations[6-8]. This review synthesizes evidence from Blue Zone populations and aging biology to identify the molecular and lifestyle factors that determine cardiovascular health.
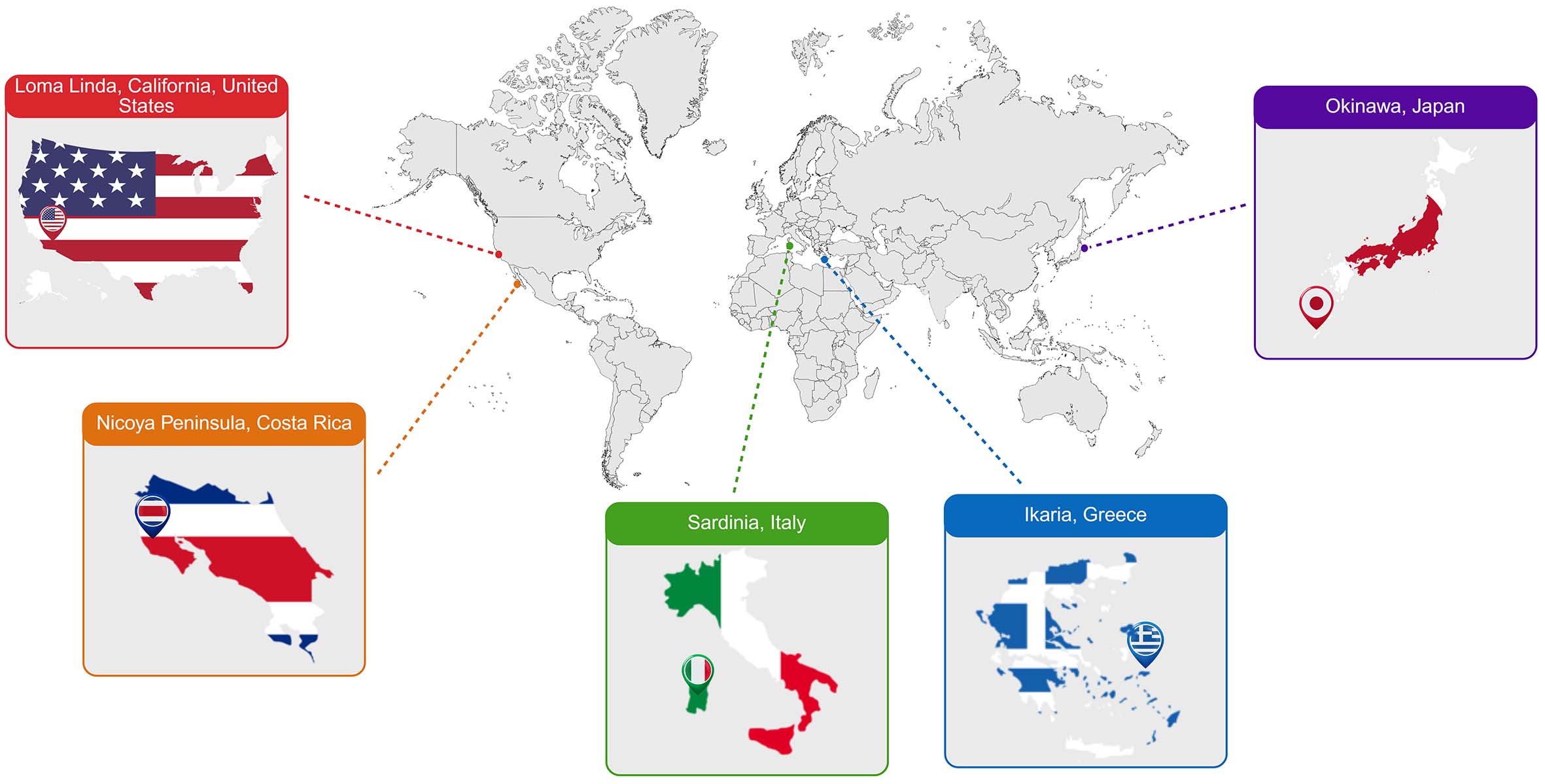
Figure 1. Geographic distribution of the Blue Zones: Okinawa, Sardinia, Ikaria, Nicoya, and Loma Linda. Created in BioRender, Javaheri, A. (2026) https://BioRender.com/jliqpcu.
Power 9 principles
To explain the lifestyle patterns underlying exceptional longevity in Blue Zones, Dan Buettner and his team identified the "Power 9" principles [Figure 2][9]. These include nine evidence-based habits observed among centenarians in these regions. The principles reflect key behaviors and social practices that are thought to contribute to extended life expectancy and improved cardiovascular and overall well-being[9].

Figure 2. The nine lifestyle principles identified across Blue Zone populations, collectively known as the Power 9. Created in BioRender, Javaheri, A. (2026) https://BioRender.com/3vti3ma.
A defining feature among these populations is the absence of structured exercise routines, which is associated with improved endothelial function, reflected by greater flow-mediated dilation (FMD)[10]. Instead, physical activity is embedded within daily life and is largely derived from habitual activities such as walking, with levels approximating at least 30 min of brisk walking on most days or 8,000-10,000 steps per day, consistent with current physical activity recommendations[11,12]. In a study of the Ikarian population, more than 90% of men and 70% of women older than 80 years reported moderate to high levels of daily activity, a pattern similarly observed among Sardinian centenarians[13-15]. Consistent with these findings, data from the Physicians’ Health Study demonstrated that individuals engaging in regular physical activity were more likely to reach the age of 90 while maintaining functional independence and a lower burden of cardiovascular disease[15]. This principle of Move Naturally appears to be fundamental to sustaining cardiovascular and metabolic health in Blue Zones[9,16].
In addition to physical activity, a clearly defined sense of purpose is a recurring theme. This is referred to as Ikigai in Okinawa and Plan de Vida in Nicoya, both of which translate to "reason for waking up in the morning." Studies suggest that having a Purpose may extend life expectancy by up to seven years[3]. A strong sense of purpose has been linked to lower cardiovascular and all-cause mortality. Meta-analytic data from over 136,000 participants across ten prospective studies demonstrated that individuals reporting a higher sense of purpose experienced significantly reduced risks of both all-cause and cardiovascular death[17].
Chronic psychological stress, a contributor to systemic inflammation and age-associated diseases, is reduced in Blue Zone communities through daily stress-reducing rituals[9]. Although these practices vary by region, such as ancestor remembrance in Okinawa, happy hour in Sardinia, afternoon napping in Ikaria, and prayer among the Adventists, they share a common purpose of lowering physiological stress[3,8,18]. This intentional stress-management approach reflects the principle of downshift[8].
Nutritional practices in Blue Zones are equally distinctive. One such practice is the 80% rule, which encourages individuals to stop eating when they feel approximately 80% full[3,19,20]. This mindful eating behavior is thought to reduce overall caloric intake and help maintain metabolic efficiency[19]. Evening meals are typically light, and more calories are consumed earlier in the day, with no food intake occurring afterward[19]. Diets are largely plant-based, emphasizing legumes such as fava beans, lentils, soy, and black beans[19]. Meat is consumed infrequently, generally no more than five times per month, and in small portions, consistent with the principle of plant slant[3,6].
Alcohol consumption, particularly in the form of wine, is moderate and socially integrated[9]. Typically, individuals consume one to two glasses per day during meals or with companions, a pattern associated with improved cardiovascular outcomes when compared to abstention or binge drinking[21,22]. Wine represents more than a source of ethanol, as it is a complex matrix rich in polyphenolic compounds[23]. Red wine contains bioactive molecules such as flavonoids and stilbenes like resveratrol that have been shown to enhance endothelial nitric oxide (NO) synthase activity, reduce low-density lipoprotein oxidation, and modulate inflammatory pathways involving IL-6 and C-reactive protein (CRP)[23]. Beyond vascular effects, red wine polyphenols appear to regulate gene networks involved in cellular stress responses and apoptosis, with emerging evidence linking multiple “wine-associated” genes to pathways regulating nitric oxide signaling, lipid metabolism, and DNA repair, consistent with the activation of longevity-related pathways[23,24]. In a Mediterranean population at high cardiovascular risk, moderate wine consumption as measured by urinary tartaric acid was associated with a lower incidence of cardiovascular disease[25]. Individuals excreting urinary tartaric acid corresponding to about 12-35 glasses of wine per month had a 50% lower risk of cardiovascular disease compared with minimal consumers (hazard ratio 0.50)[25]. Moreover, rising tartaric acid levels over time were associated with a significant risk reduction, suggesting that moderate intake may confer ongoing vascular protection[26]. Importantly, these benefits were not observed at higher consumption levels, reinforcing that cardioprotective effects are restricted to moderate wine intake[26]. Data from the PREDIMED trial also support the favorable cardiovascular outcomes of moderate wine consumption[27,28]. In a subset analysis using objective biomarkers of wine intake (urinary tartaric acid), moderate consumption was linked to up to a 50% relative reduction in major cardiovascular events, lower cardiovascular risk, and mortality[27,28]. In addition, a significant inverse relationship was observed between light-to-moderate wine consumption and the rate of myocardial infarction[27]. This aligns with the principle of wine at 5[9].
Social networks play an important role in maintaining long-term health[29,30]. In Okinawa, lifelong social groups known as moais provide continuous emotional support, reinforcing healthy behaviors and buffering against loneliness, an example of the right tribe principle, where individuals are embedded within communities that promote well-being[9]. This cultural practice aligns with large-scale epidemiologic evidence showing that strong social connections confer a 50% higher likelihood of survival, an effect comparable to quitting smoking and greater than reducing major risk factors such as obesity or physical inactivity[31]. These findings highlight that meaningful social integration is not only culturally embedded in Blue Zones but also a powerful biological determinant of healthy aging[32].
Religious participation is another contributing factor. Regular attendance at faith-based services has been associated with a significant increase in life expectancy, with some reports citing gains of up to fourteen years[33]. Higher levels of religiosity and spirituality were associated with more favorable cardiovascular health behaviors and overall risk profiles in African Americans[34]. Greater religious attendance was linked to increased odds (OR [95%CI]) of achieving intermediate/ideal levels of physical activity (1.16 [1.06-1.26]), diet (1.10 [1.01-1.20]), smoking (1.50 [1.34-1.68]), blood pressure (1.12 [1.01-1.24]), and overall Life’s Simple 7 (LS7) score (1.15 [1.06-1.26])[34]. Private prayer was associated with improved diet (1.12 [1.03-1.22]) and smoking (1.24 [1.12-1.39]), while religious coping showed similar associations with physical activity (1.18 [1.08-1.28]), diet (1.10 [1.01-1.20]), smoking (1.32 [1.18-1.48]), and LS7 score (1.14 [1.04-1.24]), these relationships remained significant after adjustment for stress and social factors[34]. In addition, religious beliefs and values were significantly associated with lower CRP levels (β: -0.05, 95%[CI: -0.09, -0.01])[35]. Regular participation in faith-based activities has been associated with about one-third lower all-cause mortality, a benefit comparable to several medical interventions[36]. In the Nurses’ Health Study, women who attended services weekly showed significantly lower mortality from cardiovascular disease, cancer, and all causes[37].
A strong emphasis is placed on family and intergenerational cohesion[4]. Older adults often live with or near their children and grandchildren, receive care from relatives, and remain actively involved in family life, maintaining close familial relationships, committing to long-term partnerships, and investing time in loved ones embodies the principle of Loved Ones First, which supports emotional and physical resilience across the lifespan[3]. This could be an important contributor to their longevity, since socially isolated adults are at an increased risk of premature mortality[38,39]. Among older women, loneliness was linked to an increased risk of cardiovascular mortality (HR 2.25, 95%CI: 1.14-4.45), and this association persisted after multivariable adjustment (HR 2.42, 95%CI: 1.04-5.65), supporting loneliness as an independent predictor of cardiovascular death[40]. Moreover, higher levels of loneliness or social isolation were associated with an increased risk of cardiovascular disease (HR 1.16, 95%CI: 1.1-1.22), corresponding to a 16% increase in risk (P < 0.0001)[41]. Further, Loneliness is associated with elevated levels of interleukin-6 (IL-6), whereas social isolation is linked to increased concentrations of CRP and fibrinogen, supporting a role for social disconnection in promoting systemic inflammation[42,43]. Elevated CRP levels were also associated with loneliness in older adults aged 70-90 years (β = 0.16, P = 0.02)[44]. Social isolation was also consistently associated with elevated levels of soluble urokinase plasminogen activation receptor (suPAR), a marker of chronic systemic inflammation[45]. Across multiple cohorts, suPAR shows a more robust association with social isolation than traditional acute-phase reactants such as CRP and IL-6, indicating that it more accurately reflects the cumulative inflammatory burden of long-term social disconnection[45]. Higher suPAR levels are present in heart failure (HF) and predict adverse outcomes, while in individuals without HF, they independently predict incident HF[46,47]. As a marker of immune activation, suPAR reflects underlying processes such as oxidative stress and inflammation that contribute to disease development[46].
The Blue Zones
Okinawa stands out in Japan because its exceptional longevity arises not from socioeconomic advantage, but from a distinct lifestyle and food culture shaped by its subtropical geography, Ryukyu heritage, and a resource-efficient diet[48,49]. Despite being Japan’s poorest prefecture, Okinawa has a higher life expectancy than the national average[50]. Population in Okinawa traditionally followed a low-calorie, nutrient-dense eating pattern built around sweet potatoes, seaweed (kombu, mozuku), tofu, and mixed plant-animal dishes that incorporate pork in collagen-rich cuts, all complemented by very low salt intake and mineral-rich drinking water from coral-derived limestone aquifers[6]. Additionally, their diet includes substantial collagen intake from pork cartilage and tendon, as well as iodine-rich Kombu seaweed and bitter gourd (Goya), which has been associated with hypoglycemic effects[6]. Moreover, their overall caloric intake remains low, a factor believed to contribute to their extended health span[6,51-53]. These foods provide bioactive compounds linked to lower cardiovascular and metabolic risk, including n-3 fatty acids, fucoidan, elastin- and collagen-derived amino acids, and phytochemicals such as momordin[53]. Cultural norms around hara hachi bu (stopping at 80% fullness), strong social cohesion, and multigenerational support networks buffer chronic disease risk, consistent with cohort studies showing that diet quality, social integration, and physical activity patterns explain most of Okinawa’s advantage over socioeconomic predictors of mortality[6,50].
Sardinia's unusually high concentration of male centenarians is linked to a combination of geographic isolation, genetic homogeneity, traditional dietary patterns, and deeply rooted social structures[54]. The inland mountain regions, particularly the Ogliastra and Barbagia areas, historically maintained a pastoral lifestyle characterized by regular low-intensity physical activity, modest caloric intake, and diets centered on whole grains, legumes, garden vegetables, sheep and goat dairy products rich in omega-3 and conjugated Linoleic acid, and limited meat consumption[54]. Members of those communities regularly walk five or more miles per day over mountainous terrain[3]. Studies highlight the region’s exceptionally strong family cohesion, high social integration, and low levels of chronic psychosocial stress as additional factors contributing to reduced cardiovascular and metabolic disease risk[55,56]. Genetic analyses have identified enriched frequencies of certain alleles associated with inflammation modulation and lipid metabolism, although lifestyle factors explain most observed longevity advantages[57]. Overall, Sardinia illustrates how the interplay of environment, culture, and consistent lifelong behaviors can produce a population with both an extended lifespan and comparatively low age-related morbidity[58].
Ikaria’s longevity profile is rooted in a combination of Mediterranean dietary traditions, sustained daily physical activity, and unusually strong social cohesion that collectively reduce cardiometabolic and inflammatory burden across the lifespan[8,15]. The island’s traditional diet emphasizes legumes, wild leafy greens, potatoes, moderate fish intake, minimal processed foods, and generous use of olive oil, with herbal infusions rich in polyphenols contributing additional antioxidant and vasodilatory effects[59-62]. Epidemiological studies note low rates of hypertension and cardiovascular mortality relative to mainland Greece, attributed not only to dietary patterns but also to habitual walking in mountainous terrain and daily routines that integrate natural movement[14,63]. Social structures characterized by dense family networks, communal gatherings, and low levels of chronic stress appear to protect against frailty and cognitive decline[64,65]. Sleep patterns, including widespread afternoon napping and moderate consumption of locally produced wine, have also been associated with healthier cardiometabolic profiles[8]. Favorable sleep habits may also contribute to the island’s cardiometabolic profile, with studies reporting generally good sleep quality and an average sleep duration of approximately 9 hours per night among Ikarian adults[66]. Good sleep quality and regular sleep patterns have been associated with improved cardiometabolic health, lower risk of metabolic syndrome and type 2 diabetes, and reduced risk of atherosclerotic cardiovascular disease[67-69]. Together, these lifestyle and environmental features create a synergistic context in which chronic disease risk is consistently attenuated, supporting the island’s high proportion of healthy older adults[14].
Nicoya in Costa Rica exhibits remarkable longevity attributed to a combination of dietary quality, environmental conditions, and strong social structures[3]. Residents traditionally consume a Mesoamerican diet centered on beans, corn tortillas, squash, tropical fruits, and limited animal protein, providing high fiber, low glycemic load, and favorable micronutrient density, particularly calcium and magnesium from water sources filtered through limestone aquifers[6,70,71]. Epidemiological studies report lower rates of cardiovascular mortality and metabolic disease, partly linked to low overall caloric intake, sustained physical activity through agricultural and household labor, and preservation of lean body mass into older adulthood[72,73]. Strong family networks, high intergenerational support, and cultural emphasis on purpose and community engagement contribute to lower chronic stress and improved healthy aging trajectories[74]. These factors create a stable physiological environment in which inflammation, frailty, and cardiometabolic decline progress more slowly than in comparable populations[72,73].
Loma Linda, California, is distinguished by its large Seventh-day Adventist population, whose health-related behaviors contribute to some of the longest life expectancies in North America[3,75]. Epidemiological analyses from the Adventist Health Studies demonstrate significantly lower incidence of cardiovascular disease, hypertension, type 2 diabetes, and certain cancers, largely attributed to plant-forward dietary patterns that emphasize whole grains, legumes, nuts, fruits, and vegetables, alongside minimal intake of processed foods and red meat[5,76]. Additional factors include abstention from tobacco and alcohol, regular moderate physical activity, consistent sleep routines, and strong participation in faith-based social networks that enhance emotional support and reduce chronic stress[77,78]. These lifestyle practices produce favorable lipid profiles, lower body mass index, reduced systemic inflammation, and improved metabolic control, contributing to exceptional longevity and lower age-adjusted morbidity compared with national averages[78-80].
Cardiovascular phenotype of Blue Zones
Epidemiologic data from Blue Zones demonstrate reduced cardiovascular risk compared with surrounding populations. In Ikaria, Greece, the incidence of stroke and heart disease is approximately 50% lower than in the rest of the country[63]. Similarly, in Sardinia, Italy, the recurrence of cerebrovascular events is lower, and Italians outside Sardinia are about 30% more likely to develop coronary heart disease[81]. Okinawa, Japan, is recognized as one of the regions with the lowest prevalence of heart disease worldwide, with Americans exhibiting nearly 80% higher risk compared with Okinawans[82]. In Nicoya, Costa Rica, diets rich in white rice and beans, together with drinking water high in calcium and magnesium, have been associated with reduced cardiovascular mortality[6]. Furthermore, studies from Loma Linda University show that Seventh-day Adventist men have about 40%-50% lower mortality from ischemic heart disease, largely due to their non-smoking, predominantly vegetarian lifestyle and other healthy habits, rather than genetic differences[79]. These advantages point toward a shared biological architecture of cardiovascular protection, which can be understood by examining three interrelated domains that emerge across Blue Zone research:
1. Vascular aging and endothelial function
Populations in Blue Zones exhibit vascular aging profiles that are markedly different from those of typical high-income populations, characterized by preserved endothelial function, lower arterial stiffness, and reduced age-related vascular impairment[19]. Much of this effect maps to habitual regular low-intensity physical activity and dietary patterns that improve endothelial reactivity and slow pulse-wave velocity progression[83]:
(i) Diet: Plant-based diets rich in polyphenols (e.g., Ikarian olive oil, Sardinian red wine), n-3 fatty acids, and low sodium[84,85]. Minimal processed foods, with > 90% home-prepared meals emphasizing legumes, tubers, and antioxidant-rich ingredients like Ikarian herbal tea (anti-inflammatory) and Sardinian coffee[14,86].
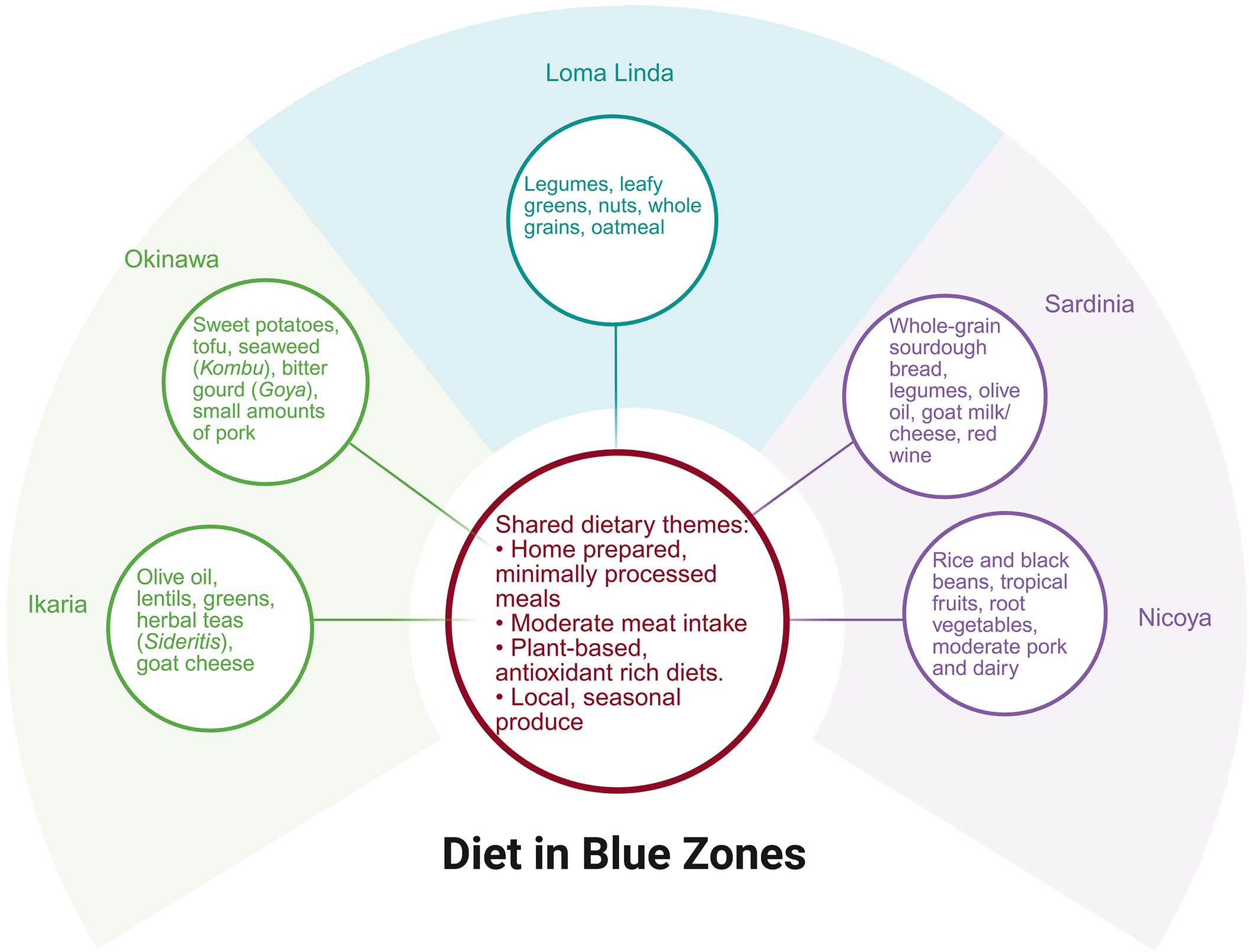
Figure 3. Summary of dietary patterns across the five Blue Zones. Created in BioRender,Javaheri, A. (2026) https://BioRender.com/r7x0gd5.
Many studies conclude that Mediterranean-style and low-calorie diets, dietary patterns closely resembling those observed in Blue Zones, enhance endothelial function and improve surrogate markers of vascular aging, offering a plausible mechanistic explanation for the low incidence of cardiovascular disease in these populations[87,88]. For instance, in a randomized controlled trial of individuals with high cardiometabolic risk, adherence to a Mediterranean diet significantly improved endothelial function, with FMD increasing from 6.1% to 9.8% in the fasting state over 8 weeks, compared with 5.0% in controls (P = 0.008)[89]. Postprandial endothelial responses were also markedly enhanced, with FMD reaching 8.99% vs. 3.86% in the control group (P = 0.003), indicating both acute and sustained vascular benefits[89]. In line with these findings, a meta-analysis of 21 independent datasets, which demonstrated a significant overall improvement in endothelial function with the Mediterranean diet (Standard mean difference: 0.34; P = 0.0001), including an approximate 1.39% increase in FMD (P < 0.001), with consistent effects observed across both healthy individuals and those at increased cardiovascular risk, and independent of age, body mass index, or study duration[90]. Further, the Mediterranean diet, when compared to a low-fat diet, was associated with modest increases in HDL cholesterol (+0.76 mg/dL; P = 0.02) and reductions in fasting blood glucose (-0.96 mg/dL;
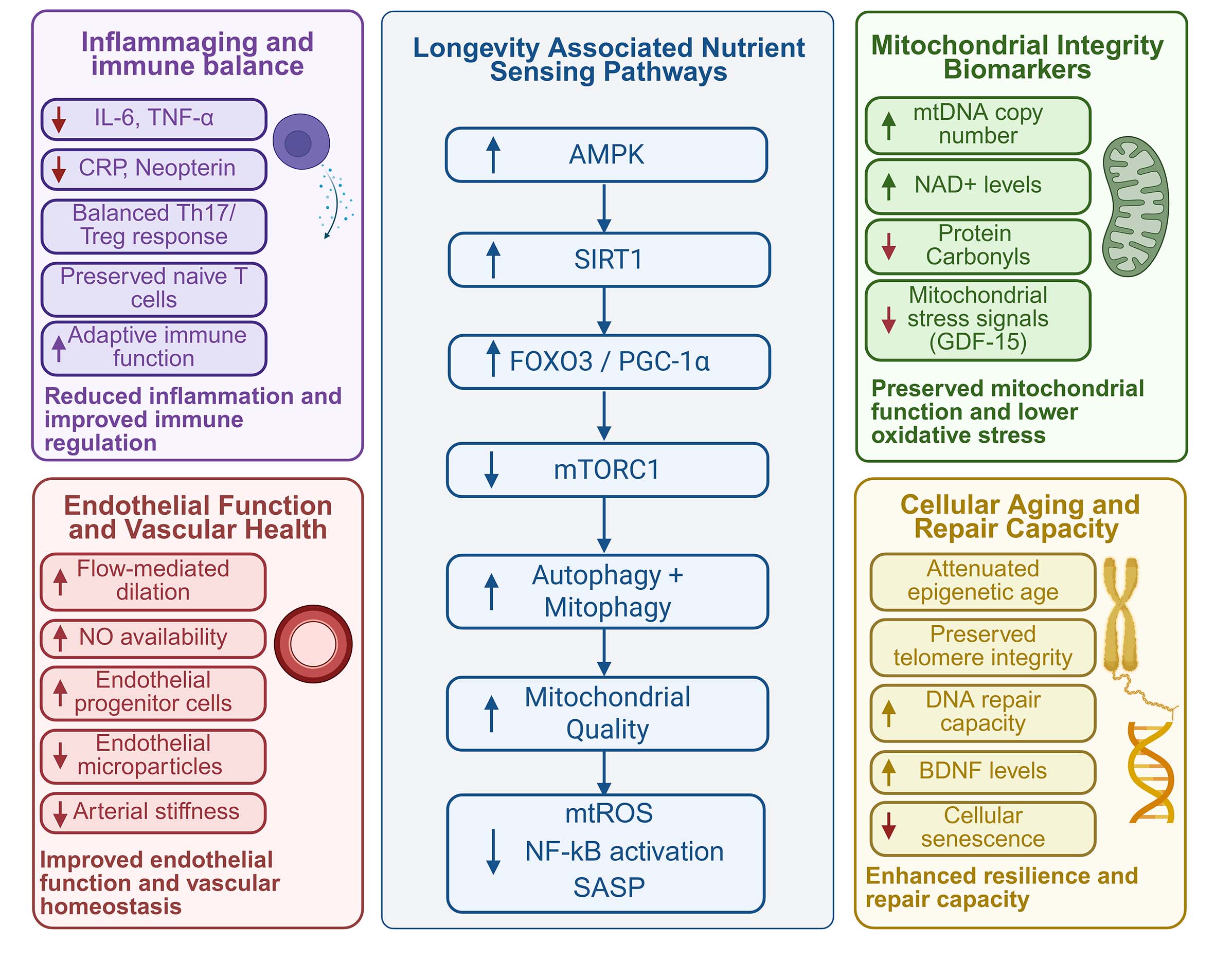
Figure 4. Proposed mechanistic framework linking Blue Zone lifestyle patterns to healthy aging and longevity. Created in BioRender, Javaheri, A. (2026) https://BioRender.com/wmhm7he.
(ii) Physical activity: Physical activity that raises heart and respiratory rates, including walking, running, cycling, and similar movements, forms the basis of aerobic exercise[97]. The 2022 World Health Organization guidelines recommend 150-300 min of moderate-intensity activity (e.g., walking, cycling, ballroom dancing) or 75-150 min of vigorous-intensity activity (e.g., jogging, swimming, hiking) per week to achieve optimal health outcomes[98-100]. Aerobic exercise induces central and peripheral cardiovascular adaptations such as enhanced myocardial contractility, increased ventricular filling, and improved maximal cardiac output, while lowering blood pressure in hypertensive patients[101,102].
Regular activity also improves lipid profiles, reduces thromboembolic complications in atrial fibrillation, supports weight management, increases insulin sensitivity, lowers HbA1c, and reduces systemic inflammation, partly reflected in decreased CRP and improved autonomic function[103-105]. The molecular mechanisms underlying the protective effects of exercise on human health are diverse[104]. Intensive exercise programs increase circulating energy substrates such as free fatty acids and glucose, which are subsequently taken up by skeletal muscle cells[104]. Repeated bouts of exercise enhance the uptake and oxidation of these substrates, a process mediated by signaling pathways including adenosine monophosphate-activated protein kinase (AMPK) and β-adrenergic signaling[104,106-108]. Exercise also activates pathways that regulate mitochondrial biogenesis in skeletal muscle, such as NO synthase, sirtuins, and peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α), and strengthens endothelial integrity through nitric oxide-mediated anti-apoptotic signaling and adaptive mitohormesis[109-115]. Exercise additionally attenuates leukocyte-driven inflammation by inhibiting leptin-mediated hematopoietic activation[116]. At the cardiac level, it supports favorable left ventricular remodeling after myocardial infarction and reduces NT-proBNP[117,118]. Emerging work on exercise-induced “exerkines” suggests roles for musclin, myonectin, interleukin-8, and fibroblast growth factors in blood pressure reduction and angiogenesis. At the same time, skeletal muscle-derived IL-6 contributes to anti-inflammatory and cardioprotective effects[119,120]. Exercise also induces beneficial epigenetic changes, including PGC-1α upregulation, which may delay complications of type 2 diabetes[121,122].
Quantitative analysis of physical activity in Blue Zone populations indicates that activity is predominantly low-to-moderate in intensity and sustained throughout the day, rather than performed in discrete exercise sessions[123]. Across studies, approximately 60%-80% of reported activities are classified as moderate intensity, with regional variation[14,66,124]. In comparison, the Okinawan population exhibits a higher proportion of low-intensity activity (83%), reflecting differences in cultural and occupational patterns[123,125]. In Sardinian cohorts, older adults (≥ 90 years) accumulate approximately 12,110 steps/day in men and 12,799 steps/day in women, while shepherd populations walk a mean distance of 12.4 ± 7.8 km/day, often across steep terrain (average slope of 15%), representing sustained moderate-intensity exertion maintained throughout the year[123,126,127].
(iii) Mitochondrial integrity: Blue Zone dietary patterns may attenuate vascular aging, in part through mechanisms known to support mitochondrial integrity, a central determinant of endothelial function[84,128,129]. Mitochondrial dysfunction, characterized by elevated mitochondrial ROS, impaired biogenesis, disrupted fission-fusion balance, and defective mitophagy, plays a major role in endothelial impairment, reducing NO bioavailability and promoting inflammation, apoptosis, and arterial stiffening[130-132]. Because endothelial cells depend heavily on mitochondrial redox control, maintaining mitochondrial quality is essential for vascular homeostasis and NO signaling[133,134]. Interventions that enhance mitochondrial efficiency or limit mtROS, including AMPK activation, SIRT1 signaling, and improved metabolism, have been shown to preserve endothelial function and slow age-related vascular decline[135,136] [Figure 4]. These mechanisms align with dietary characteristics common in Blue Zones, where plant-forward, calorie-light eating patterns may reduce oxidative burden, stabilize glycemia, and promote mitophagy-driven mitochondrial renewal[137]. Experimental evidence shows that improving mitochondrial quality control protects the vasculature from oxidative injury and endothelial dysfunction, positioning mitochondria as the central hub linking metabolic lifestyle factors with vascular aging[138-141].
Accumulating evidence indicates that aging is accompanied by measurable alterations in circulating biomarkers reflecting mitochondrial stress and oxidative damage[142,143]. Among these, growth differentiation factor-15 (GDF-15) has emerged as a robust marker of mitochondrial dysfunction, showing strong associations with the pace of biological aging, epigenetic aging indices, and age-related conditions such as sarcopenia and mortality[142,144]. Elevated GDF-15 levels appear to reflect early mitochondrial stress responses that precede overt tissue damage, suggesting that mitochondrial dysregulation may represent an upstream driver of the aging process[142]. In contrast, markers of oxidative stress, including allantoin and protein carbonyls, provide insight into the downstream consequences of mitochondrial dysfunction[142]. Allantoin has been associated with accelerated aging trajectories and phenotypic aging measures, whereas protein carbonyls reflect cumulative oxidative damage[142].
In human centenarian cohorts, these processes can be evaluated using functional assays and circulating biomarkers[145]. Mitochondrial biogenesis, assessed through oxygen consumption rate in peripheral blood mononuclear cells, appears to be relatively preserved in centenarians, while mitochondrial DNA copy number serves as a proxy for mitochondrial integrity[145] [Figure 4]. At the same time, increased oxidative stress, reflected by higher levels of protein carbonylation, is associated with poorer cognitive function, whereas enhanced DNA repair capacity, indicated by higher activity of the base excision repair enzyme APE1, correlates with better cognitive outcomes[145,146]. Circulating factors such as brain-derived neurotrophic factor (BDNF) and NAD+ levels are associated with preserved cognitive function, highlighting mitochondrial integrity, oxidative balance, and repair capacity as key drivers of functional aging[145].
2. Inflammation, oxidative stress, and metabolic health
Blue Zones exhibit attenuated chronic inflammation (IL-6, CRP) and favorable metabolic markers compared with matched non-Blue Zone populations, despite some heterogeneity across sites. The improvement in metabolic health originates mainly from:
(i) Antioxidant synergy: Diets high in fiber with phytonutrients, whole grains, vegetables, and omega-3 fatty acids (Okinawan seaweed/iodine, Loma Linda nuts/lignans) suggest a 40% reduction in cardiovascular mortality[147-149]. These nutrients reduce circulating IL-6, CRP, and Tumor Necrosis Factor-α signaling, attenuating inflammation-derived aging and oxidative stress[150-152].
(ii) Metabolic flexibility: Several Blue Zone populations characteristically consume larger meals earlier in the day, followed by lighter evening intake with minimal or no subsequent food consumption[3]. Increasing evidence indicates that restricting caloric intake to daytime hours and minimizing nocturnal eating may improve insulin sensitivity, glucose homeostasis, oxidative stress, adiposity, and overall cardiometabolic function by reducing circadian misalignment between feeding behavior and endogenous metabolic rhythms[153-155]. At the molecular level, these dietary patterns, such as time-restricted eating and polyphenol-rich foods, may converge on nutrient and stress-sensing pathways, particularly AMPK-SIRT1-FOXO signaling[156,157] [Figure 4]. AMPK is activated during energy depletion and promotes catabolic metabolism, fatty acid oxidation, mitochondrial biogenesis, and inhibition of anabolic mTOR signaling[157-159]. AMPK activation can also increase NAD+ availability and enhance SIRT1 activity, thereby linking cellular energy status to transcriptional programs involved in stress resistance and mitochondrial adaptation[157]. SIRT1 deacetylates FOXO transcription factors, including FOXO1 and FOXO3, enhancing the expression of antioxidant and stress-response genes such as superoxide dismutase, catalase, and glutathione-related enzymes[157,160]. Through downstream effects on PGC-1α-mediated mitochondrial biogenesis, autophagy, mitophagy, ROS detoxification, and suppression of NF-κB/SASP (senescence-associated secretory phenotype) related inflammatory signaling, AMPK-SIRT1-FOXO signaling provides a plausible link between Blue Zone dietary patterns and preserved metabolic flexibility[157,161]. Activation of this pathway may enhance insulin sensitivity, improve lipid handling, reduce ectopic fat accumulation, and preserve mitochondrial efficiency, thereby supporting cardiometabolic resilience with aging[157]. By limiting oxidative damage and chronic low-grade inflammation, AMPK-SIRT1-FOXO signaling may also contribute to a lower risk of age-related metabolic dysfunction, cardiovascular disease, and functional decline[157].
(iii) Branched-chain amino acids (BCAAs) balance: BCAAs, particularly leucine, add another layer to metabolic health by modulating nutrient-sensing pathways that intersect cellular growth and stress pathways such as the mammalian target of rapamycin (mTOR)[162-164]. Moderate (not high) total protein intake, combined with sufficient BCAAs from plant sources such as legumes and whole grains, delivers high-quality protein while reducing glycemic load and the cardiometabolic risk associated with high animal-protein and high-insulinogenic diets[5,165-168]. Animal protein from additive-free, locally raised pork/goat (Okinawa, Sardinia) or dairy (Nicoya) ensures high quality without metabolic stress[54,70]. Recent multi-organ proteomics show that moderate protein restriction in midlife reprograms the aging proteome across dozens of tissues and attenuates age-associated molecular signatures, with particularly strong benefits in adipose and cardiovascular tissues consistent with diminished activation of growth pathways, including those downstream of mTOR[169]. Conversely, BCAAs retain clinical value for preserving lean mass and functional reserve in older adults or chronically ill patients, so the goal is recalibration rather than elimination[170]. Intriguingly, pharmacologic modulation of nutrient-sensing can phenocopy some dietary effects[171]. For instance, low-dose glucagon-like peptide-1 receptor agonists (GLP-1RAs) produce broad, body-wide multi-omic “age-counteracting” signatures and improved muscle/functional readouts in aged mice, and those molecular signatures closely resemble the effects of mTOR inhibition (rapamycin)[171]. These lines of evidence argue for an unresolved tension in the field. Long-lived populations in the Blue Zones skew toward modest to low protein intake, with predominantly plant-derived amino acids[172]. However, confirming studies in controlled human settings are still needed to clarify whether this pattern reflects causal biology or cultural confounders[173-176].
CELLULAR AND MOLECULAR AGING
At the cellular level, evidence from centenarian and long-lived cohorts suggests slower accrual of molecular aging signatures in Blue Zones:
(i) Telomere/epigenetic preservation: Emerging evidence from both interventional and observational studies supports the notion of preserved telomere stability and slow epigenetic aging[177]. Blue Zones consistently obtain polyphenols from region-specific foods, including herbs and mountain teas (Ikaria), Cannonau wine and olive oil (Sardinia), turmeric and cruciferous vegetables (Okinawa), beans and squash (Nicoya), and nuts, berries, and legumes (Loma Linda)[84]. These polyphenol-enriched diets are associated with reductions in epigenetic age, as measured by multiple DNA methylation clocks, in randomized controlled trials in humans[178]. In the DIRECT PLUS trial, adherence to a Green-Mediterranean diet enriched with polyphenol-containing foods was associated with an attenuation of DNA methylation age (mAge)[178]. Higher urinary levels of specific polyphenol metabolites, including hydroxytyrosol, urolithin C, and tyrosol, were inversely correlated with 18-month changes in mAge, with tyrosol remaining independently associated after adjustment for age, sex, and weight loss[178]. These changes were further linked to differential methylation at specific aging-related CpG sites like cg16290275, which supports a potential epigenetic mechanism[178]. Meanwhile, caloric restriction in animal models has been shown to decelerate age-related methylation changes across tissues[179]. Time-restricted eating (Sardinia, Loma Linda) and religious fasts (Ikaria, Adventists) yield metabolic benefits, including improved insulin sensitivity, reduced fat mass, and reductions in blood pressure, all of which are major contributors to cardiovascular events[180]. One key benefit of intermittent nutrient restriction is its effect on autophagic flux by inhibiting mTOR, thereby enhancing autophagy, a cellular recycling process that degrades damaged proteins and organelles to maintain cellular homeostasis[181-183]. Many of the beneficial effects of caloric restriction are thought to involve suppression of nutrient-sensing pathways, particularly mTORC1 signaling[184]. The observation that lifespan extension by caloric restriction is attenuated in models with pre-existing reductions in mTORC1 activity supports the presence of overlapping mechanisms[184]. Downstream, translational regulation via 4E-Binding Protein appears to play a central role, facilitating selective translation of mitochondrial proteins, enhancing mitochondrial function, and promoting proteostasis[184]. Alongside these molecular effects, improvements in muscle function have also been observed in aging mice subjected to caloric restriction[183,185]. These findings support that caloric restriction exerts its anti-aging effects, at least in part, through coordinated modulation of mTORC1 signaling and its downstream metabolic and mitochondrial pathways[184,186].
Emerging human studies also indicate that intermittent nutrient restriction, particularly through fasting-mimicking dietary approaches, is associated with measurable activation of autophagy[187-189]. In a randomized clinical trial, short-term fasting-mimicking diet intervention led to a significantly lower LC3B-II/LC3B-I ratio in peripheral blood mononuclear cells at day 6 compared with controls (0.7 ± 0.4 vs. 1.3 ± 0.7; 0.019), reflecting increased autophagic degradation and enhanced autophagic flux[188]. Further assessment using chloroquine-treated samples showed a trend toward increased autophagic flux during the intervention, with persistence following refeeding in one fasting group, suggesting sustained activation of the autophagic process[188]. In addition, fasting-based interventions have been reported to improve metabolic parameters linked to nutrient-sensing pathways, including reductions in fasting glucose, insulin resistance (HOMA-IR), and circulating IGF-1[188,190]. Increased expression of autophagy-related genes, such as GABARAPL1, ATG2A, and MAP1LC3A, has been observed in human studies, further supporting enhanced autophagic activity during caloric restriction[188,191].
In human populations, adherence to Mediterranean-style diets rich in antioxidants and fiber, and engagement in moderate physical activity, is positively associated with longer telomere length, likely by mitigating oxidative stress and systemic inflammation[192].
(ii) Improved immune cell profile: There are multiple lines of evidence that, in certain centenarian populations, immune-cell profiles reflect a more balanced inflammatory state[193-195]. For instance, one study of human centenarians found that their ratio of T helper 17 to T regulatory cells was lower than in older adults, and that their regulatory T cells secreted more anti-inflammatory cytokines, suggesting a shift in immune phenotype that might protect against chronic inflammation[196]. Further, Sardinian centenarians demonstrate a marked deviation from the expected age-associated increase in neopterin concentrations[19,197]. This pattern supports the potential utility of neopterin as a marker of survival and overall health. Importantly, the lower neopterin levels observed in centenarians align with a phenotype characterized by attenuated inflammation and oxidative stress relative to other old-old individuals[198]. Moreover, Okinawans were found to have high levels of IgG and IgA in their serum; this may be attributed to the island vegetables that they consume[199]. Increased natural killer (NK) cell activity, mediated in part by cytokine signaling pathways, such as increased IFN-γ, was observed in Okinawan residents who consume Fucoidan-derived from Cladosiphon okamuranus[200].
Centenarians across Blue Zones exhibit a coordinated immune phenotype characterized not by absence of inflammation, but by attenuated chronic inflammatory signaling coupled with preservation of immune competence[197]. In Mediterranean Blue Zones such as Sardinia and Ikaria, individuals demonstrate a modified inflammaging profile, including lower DNAm-predicted IL-6 levels, particularly in men, alongside reduced macrophage activation as reflected by decreased neopterin concentrations, suggesting reduced chronic innate immune activation[197] [Figure 4].
At the level of adaptive and regulatory immunity, centenarians display features of preserved immune “youthfulness” and refined immune control[197]. Studies from the Nicoya Peninsula in Costa Rica reveal a T-cell profile characterized by higher proportions of naïve T cells and reduced CD8+ memory T cells, indicative of delayed immunosenscence and maintained immune repertoire diversity[197,201]. This supports a model in which exceptional longevity is associated with a multi-layered immune equilibrium, encompassing reduced pro-inflammatory cytokine burden (IL-6), controlled innate immune activation (macrophage signaling), preservation of adaptive immune function (naïve versus memory T-cell balance), and optimized immune regulation (T regs)[197].
Centenarians exhibit a complex cytokine profile characterized by concurrent activation of pro- and anti- inflammatory pathways[202]. Consistent with aging, levels of pro-inflammatory cytokines, including TNF-α, IL-6, IFN-α, and IL-23, increase, along with other mediators, such as GM-CSF and osteopontin (OPN), that support a chronic inflammatory state[202]. Despite this, centenarians also demonstrate compensatory upregulation of anti-inflammatory signals, notably IL-19, a member of the IL-10 family that promotes Th2 polarization and enhances IL-10 production, potentially dampening excessive Th1-driven inflammation[202].
In contrast, cytokines regulating adaptive T-cell responses show minimal differences between centenarians and controls, suggesting that age-related T-cell dysfunction (e.g., loss of naïve T cells and accumulation of terminally differentiated cells) is not primarily driven by alterations in cytokines[202]. In the B-cell compartment, B-cell activating factor (BAFF) levels are increased, although this effect may be partially offset by elevated soluble Transmembrane activator and CAML interactor (TACI), which can neutralize BAFF activity[202]. Additional changes include increased IL-11 and bone morphogenetic proteins (BMP-4, BMP-7), which may reflect compensatory responses to age-related bone and hematopoietic decline but also contribute to immune regulation[202].
(iii) Genetic factors: Several genetic variants have demonstrated positive associations with longevity in Blue Zone populations, primarily involving pathways of stress resistance, metabolism, and immune regulation[19]. Among the most consistently reported is Forkhead/winged helix box O type 3 (FOXO3) gene, a transcription factor within the insulin/IGF-1 signaling pathway that enhances cellular resistance to oxidative stress, promotes DNA repair, and regulates apoptosis[203-206]. Specific variants, such as rs2802292, have been linked to increased lifespan, particularly in Okinawa, where carriers of the longevity-associated G-allele exhibit longer telomeres, increased telomerase activity, and age-related upregulation of FOXO3 expression[204]. In addition, this variant is associated with favorable, sex-specific modulation of inflammation, including reduced age-related increases in the pro-inflammatory cytokine IL-6 in women and increased levels of the anti-inflammatory cytokine IL-10 in men[204].
Similarly, the ApoE2 genotype has been associated with longevity and reduced risk of neurodegenerative disease and atherosclerosis (in midlife), with higher prevalence reported in some cohorts from Ikaria and Sardinia[19,207,208]. Variants in HLA class II genes (HLA-DQB1, HLA-DRB1) suggest that adaptive immune function contributes to healthy aging, potentially through modulation of chronic inflammation[19]. Additional associations have been described for genes involved in vascular and inflammatory regulation, including ACE and TNF-α, particularly in relation to cardiovascular and cognitive outcomes, while emerging evidence implicates genes such as TAS2R38, which may influence dietary behavior and metabolic responses[19,209]. Further, epidemiological evidence shows that the prevalence of G6PD (glucose-6-phosphate dehydrogenase) deficiency is very low in Nicoya, Ikaria, and the central Blue Zone region of Sardinia, suggesting that this might play a part in the longevity in Blue Zones by protecting against cellular damage from ROS species[4,210,211].
Genetics versus lifestyle
The factors underlying the development of Blue Zones remain incompletely understood and have been discussed extensively[212]. One proposed explanation is the contribution of unique genetic factors[212]. However, several investigations have not identified a clear enrichment of established longevity-associated alleles within some of these populations[213,214]. Further, twin studies from Denmark have shown that genetic factors account for approximately 20% of the variability in human lifespan, whereas the majority of longevity appears to be influenced by lifestyle and environmental factors[3]. Most studies evaluating lifestyle factors in Blue Zone populations are observational or ecological in design and therefore cannot fully disentangle lifestyle effects from genetic background, familial clustering, or population-specific ancestry[214,215]. While this does not exclude genetic contribution to exceptional longevity, it suggests that currently recognized genetic variants alone are unlikely to fully explain the remarkable lifespan observed in Blue Zones[212]. Advances in genomic technologies may yet identify rare or population-specific variants contributing to healthy aging in Blue Zone populations[212]. Nevertheless, the limited and inconsistent genetic findings to date support the view that lifestyle factors play a major role in the exceptional longevity[212]. In support of this concept, the observation that individuals who migrated out of Nicoya may lose their survival advantage is consistent with an important contribution of environmental factors beyond genetics, although this finding should be interpreted cautiously because migration studies are vulnerable to selection bias[72].
Most research has focused on older individuals within these populations, whereas the dynamic emergence of Blue Zones may offer additional insight into the sociocultural and environmental determinants of longevity[212]. Studying younger generations in Blue Zones may help identify early-life factors that promote healthy aging and survival into advanced age[212].
Limitations and potential confounding factors
The mechanistic framework presented above combine population epidemiology, randomized dietary trials, biomarker studies, and experimental aging literature. However, heterogeneity between Blue Zones (dietary components, genetic backgrounds, socioeconomic conditions) and differences in measurement methods (dietary recall, biomarker panels, epigenetic clocks) limit precise causal attribution[215]. Much of the strongest causal evidence comes from intermediary pathways (endothelial function, inflammation), where randomized trials of Mediterranean or calorie-moderation diets and exercise show reproducible benefit[83,90,91]. Still, longitudinal, multi-omic studies within Blue Zone cohorts would be required to definitively partition genetic from lifelong environmental contributions to the cardiac phenotype.
Environmental factors, like water and air quality, may confound these associations. For example, in Nicoya, drinking water is relatively rich in calcium and magnesium, which has been proposed to contribute to cardiovascular health[3,216]. Cilento, a coastal and mountainous region in southern Italy, recently recognized for increased longevity, may also illustrate the relevance of drinking water composition[217]. Municipal-level analyses of 102 communities found that areas with greater concentrations of nonagenarians and centenarians had alkaline, soft-to-moderately hard water with low sodium, favorable iron and manganese levels, and no detectable toxic heavy metals[217]. In Okinawa, the island’s coral reef-derived limestone contributes to mineral-rich drinking water, particularly higher calcium content[218,219]. In addition, air quality and climate may be relevant environmental contributors to longevity in regions such as Okinawa, Nicoya, and Cilento, which share broadly similar latitudes[215,218,220].
Socioeconomic context may also influence interpretations of Blue Zone longevity. Although higher socioeconomic status is associated with longer life expectancy, this relationship appears less consistent across Blue Zones, where many populations have historically had relatively low-income levels[215]. Okinawa, for example, has been described as an exception to the traditional social gradient, and similar patterns may apply to other Blue Zones[215]. In addition, some social and economic indicators that typically predict poorer health outcomes, including income deprivation, poverty, and unemployment, have been positively associated with the probability of reaching extreme age, despite being linked to lower survival and worse health outcomes before age 90[221].
Age validation and demographic accuracy are central methodological issues in Blue Zones research because estimates of exceptional longevity depend on reliable birth, migration, and death records[222]. Incomplete or inconsistent records may lead to misclassification of extreme age and biased estimates of centenarian prevalence or regional survival advantage[222]. Another important consideration is that Blue Zone longevity may reflect the life-course exposures of older cohorts who grew up under traditional conditions, rather than the current environment[222]. Okinawa provides an important example, as its earlier longevity advantage has declined in more recent generations, likely reflecting dietary westernization and broader modernization[222].
Healthcare access can confound longevity estimates because survival into older age depends not only on lifestyle but also on preventive care, chronic disease management, and social support systems[223,224]. For Nicoya, recent reviews note that longevity has been discussed in relation to improved socioeconomic conditions, social security, and access to free healthcare[224].
Selection bias is another important limitation in longevity studies[9,225]. Identifying regions based on a high concentration of centenarians may overestimate true longevity if the age structure of the population has been altered by migration[9,224]. For instance, out-migration of younger adults can artificially increase the proportion of very old individuals remaining in the community, while in-migration of older retirees can similarly inflate the apparent concentration of centenarians[9]. Therefore, regional clustering of centenarians should be interpreted cautiously and requires careful demographic validation, including birth records, migration histories, and population denominators, before being attributed to local lifestyle or environmental factors[224]. Survivorship bias is another limitation when studying old populations because many studies examine individuals who have already reached an advanced age[226]. These participants may represent a highly selected survivor population, making it difficult to determine whether observed lifestyle, environmental, or biological features are causal determinants of longevity or characteristics of individuals who were already resilient[226]. Cultural specificity may further limit generalizability, as Blue Zones lifestyles are closely tied to local cultural and environmental conditions and may not be directly applicable to other populations[224].
CONCLUSION
Blue Zones offer a unique population-level model of healthy aging in which lifestyle behaviors, social structures, and environmental factors appear to interact synergistically to promote cardiovascular and metabolic health. Rather than relying on a single intervention, these populations demonstrate the cumulative impact of sustained lifelong habits that align with biologic pathways involved in mitochondrial function, nutrient sensing, inflammation, endothelial health, and cellular stress resistance. The convergence of epidemiologic observations with emerging mechanistic evidence strengthens the concept that healthy aging is not solely genetically determined but is strongly shaped by modifiable behavioral and environmental exposures across the lifespan.
The translational value of the Blue Zones extends beyond longevity itself and provides actionable direction for modern public health strategies. Future interventions should prioritize creating environments that facilitate habitual physical activity through walkable communities and active transportation, while simultaneously improving access to affordable, minimally processed, plant-rich dietary patterns. Public health initiatives that raise awareness and encourage healthier meal timing, caloric moderation, and reduction of ultra-processed foods may help reduce age-related cardiovascular risk. In parallel, strengthening social connectedness through community engagement programs, intergenerational support systems, and interventions targeting loneliness and social isolation may represent an underrecognized but biologically relevant strategy to improve overall health.
At the healthcare systems level, integrating lifestyle-centered approaches into preventive cardiology and aging medicine may help shift care from reactive disease treatment toward preservation of functional health span. Future research should move beyond descriptive epidemiology toward mechanistic and implementation-focused studies capable of identifying scalable interventions that reproduce the favorable biologic profiles observed in Blue Zone populations. Longitudinal studies integrating biomarkers of inflammation, mitochondrial function, endothelial health, and biological aging may further clarify the pathways linking lifestyle patterns to cardiovascular health and longevity. Future public health strategies should also incorporate culturally adaptable and community-integrated approaches tailored to the dietary habits, social structures, and environmental contexts of diverse populations, making these changes easier to adopt and sustain within different communities. Ultimately, the Blue Zones provide not only a model of exceptional longevity but also a practical framework for extending health span and reducing the burden of age-related disease.
DECLARATIONS
Acknowledgements
The graphical abstract was created with BioRender, Javaheri, A. (2026) https://BioRender.com/v2fj96v.
Authors’ contributions
Wrote the first draft of the manuscript: Imam A, Javaheri A
Provided critical revision to the manuscript: Rashidi O, Apte ART, Ataran A, Franco A
Created the figures using BioRender: Imam A
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool Grammarly (version 1.2.268.1900, released 2026-05-06) was used solely for language editing. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
Javaheri A was supported by K08HL138262 and 1R01HL155344 from the National Heart, Lung, and Blood Institute and by the Children’s Discovery Institute of Washington University and St Louis Children’s Hospital (MC-FR-2020-919), as well as the Diabetes Research Center at Washington University in St Louis of the National Institutes of Health (NIH) under award P30DK020579 and NIH grant P30DK056341 (Nutrition Obesity Research Center) and received support from the Longer Life Foundation and contract support from Bitterroot Bio.
Conflict of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Gianfredi V, Nucci D, Pennisi F, Maggi S, Veronese N, Soysal P. Aging, longevity, and healthy aging: the public health approach. Aging Clin Exp Res. 2025;37:125.
2. World population ageing; 2019. 37p. Available from: https://digitallibrary.un.org/record/3846855?ln=en&v=pdf [Last accessed on 15 Jul 2026].
3. Buettner D, Skemp S. Blue zones: lessons from the world’s longest lived. Am J Lifestyle Med. 2016;10:318-21.
5. Liu T, Gatto NM, Chen Z, et al. Vegetarian diets, circulating miRNA expression and healthspan in subjects living in the Blue Zone. Precis Clin Med. 2020;3:245-59.
6. Pes GM, Dore MP, Tsofliou F, Poulain M. Diet and longevity in the blue zones: a set-and-forget issue? Maturitas. 2022;164:31-7.
7. Zábó V, Csiszar A, Ungvari Z, Purebl G. Psychological resilience and competence: key promoters of successful aging and flourishing in late life. GeroScience. 2023;45:3045-58.
8. Legrand R, Nuemi G, Poulain M, Manckoundia P. Description of lifestyle, including social life, diet and physical activity, of people ≥ 90 years living in Ikaria, a longevity blue zone. Int J Environ Res Public Health. 2021;18:6602.
9. Buettner D. Lessons from the blue zones: there is no silver bullet (or magic pill) for a long, healthy life. Am J Lifestyle Med. 2025;19:1052-62.
10. Siasos G, Chrysohoou C, Tousoulis D, et al. The impact of physical activity on endothelial function in middle-aged and elderly subjects: The Ikaria study. Hellenic J Cardiol. 2013;54:94-101.
11. Ungvari Z, Fazekas-Pongor V, Csiszar A, Kunutsor SK. The multifaceted benefits of walking for healthy aging: from blue zones to molecular mechanisms. GeroScience. 2023;45:3211-39.
12. Piercy KL, Troiano RP, Ballard RM, et al. The physical activity guidelines for Americans. JAMA. 2018;320:2020.
13. Pes G, Tolu F, Poulain M, et al. Lifestyle and nutrition related to male longevity in Sardinia: an ecological study. Nutr Metab Cardiovasc Dis. 2013;23:212-9.
14. Panagiotakos DB, Chrysohoou C, Siasos G, et al. Sociodemographic and lifestyle statistics of oldest old people (>80 years) living in ikaria island: the ikaria study. Cardiol Res Pract. 2011;2011:679187.
15. Pietri P, Stefanadis C. Cardiovascular aging and longevity: JACC state-of-the-art review. J Am Coll Cardiol. 2021;77:189-204.
16. Buettner D. The Blue Zones Solution; New York, 2015. Available from: https://books.google.com/books?hl=en&lr=&id=cW9aBAAAQBAJ&oi=fnd&pg=PA13&ots=1AudwyXKh2&sig=z2nEmTser18IxWD7lXxjlUNIfVQ#v=onepage&q&f=false [Last accessed on 15 Jul 2026].
17. Cohen R, Bavishi C, Rozanski A. Purpose in life and its relationship to all-cause mortality and cardiovascular events: a meta-analysis. Psychosom Med. 2016;78:122-33.
18. Suzuki T. Uncollected bones and ambiguous loss: okinawan mourning rituals in the Northern Mariana Islands. Omega. 2020;84:1175-92.
19. Kreouzi M, Theodorakis N, Constantinou C. Lessons learned from blue zones, lifestyle medicine pillars and beyond: an update on the contributions of behavior and genetics to wellbeing and longevity. Am J Lifestyle Med. 2022;18:750-65.
20. Shirai T, Tsushita K. Lifestyle medicine and japan’s longevity miracle. Am J Lifestyle Med. 2024;18:598-607.
21. Van Den Brandt PA, Brandts L. Alcohol consumption in later life and reaching longevity: the Netherlands Cohort Study. Age Ageing. 2020;49:395-402.
22. Streppel MT, Ocké MC, Boshuizen HC, Kok FJ, Kromhout D. Long-term wine consumption is related to cardiovascular mortality and life expectancy independently of moderate alcohol intake: the Zutphen Study. J Epidemiol Community Health. 2009;63:534-40.
23. Gajos G. Wine and cardiometabolic risk: the view of a cardiologist and a sommelier. Eur Heart J. 2026;47:2017-9.
24. Lacayo P, Martignoni A, Park K, Castro C, Murakami S. Red-Wine Gene Networks Linked to Exceptional Longevity in Humans. Biomolecules. 2025;15:1414.
25. Lucerón-Lucas-Torres M, Saz-Lara A, Díez-Fernández A, et al. Association between wine consumption with cardiovascular disease and cardiovascular mortality: a systematic review and meta-analysis. Nutrients. 2023;15:2785.
26. De Gaetano G, Costanzo S, Di Castelnuovo A. Wine consumption and cardiovascular health: the unresolved French paradox and the promise of objective biomarkers. Eur Heart J. 2025;46:173-5.
27. Martínez-González MA, Bes-Rastrollo M, Sayon-Orea C, et al. Wine consumption, Mediterranean diet, and cardiovascular risk in two Spanish cohorts. Eur Heart J. 2026:ehaf1081.
28. Domínguez-López I, Lamuela-Raventós RM, Razquin C, et al. Urinary tartaric acid as a biomarker of wine consumption and cardiovascular risk: the PREDIMED trial. Eur Heart J. 2025;46:161-72.
29. Tan L, Hilliard A, Peverini R, et al. Navigating the Healthcare Conundrum: Leadership Perspective from a Premier Healthcare Organization in Loma Linda's Blue Zone. J Healthc Leadersh. 2024;16:83-91.
30. Marston HR, Niles-Yokum K, Silva PA. A commentary on blue zones: a critical review of age-friendly environments in the 21st century and beyond. Int J Environ Res Public Health. 2021;18:837.
31. Holt-Lunstad J. Social connection as a critical factor for mental and physical health: evidence, trends, challenges, and future implications. World Psychiatry. 2024;23:312-32.
32. Holt-Lunstad J, Smith TB, Layton JB. Social relationships and mortality risk: a meta-analytic review. PLoS Med. 2010;7:e1000316.
33. Chen Y, Kim ES, Vanderweele TJ. Religious-service attendance and subsequent health and well-being throughout adulthood: evidence from three prospective cohorts. Int J Epidemiol. 2020;49:2030-40.
34. Brewer LC, Bowie J, Slusser JP, et al. Religiosity/Spirituality and cardiovascular health: the American Heart Association life's simple 7 in African Americans of the jackson heart study. J Am Heart Assoc. 2022;11:e024974.
35. Britt KC, Boateng AC, Sebu J, et al. The association between religious beliefs and values with inflammation among Middle-age and older adults. Aging Mental Health. 2024;28:1343-50.
36. Dominguez LJ, Veronese N, Barbagallo M. The link between spirituality and longevity. Aging Clin Exp Res. 2024;36:32.
37. Li S, Stampfer MJ, Williams DR, Vanderweele TJ. Association of religious service attendance with mortality among women. JAMA Intern Med. 2016;176:777.
38. Hodgson S, Watts I, Fraser S, Roderick P, Dambha-Miller H. Loneliness, social isolation, cardiovascular disease and mortality: a synthesis of the literature and conceptual framework. J R Soc Med. 2020;113:185-92.
39. Golaszewski NM, Lacroix AZ, Godino JG, et al. Evaluation of social isolation, loneliness, and cardiovascular disease among older women in the US. JAMA Netw Open. 2022;5:e2146461.
40. Novak M, Waern M, Johansson L, et al. Cardiovascular and all-cause mortality attributable to loneliness in older Swedish men and women. BMC Geriatr. 2020;20:201.
41. Albasheer O, Abdelwahab SI, Zaino MR, et al. The impact of social isolation and loneliness on cardiovascular disease risk factors: a systematic review, meta-analysis, and bibliometric investigation. Sci Rep. 2024;14:12871.
42. Smith KJ, Gavey S, Riddell NE, Kontari P, Victor C. The association between loneliness, social isolation and inflammation: a systematic review and meta-analysis. Neurosci Biobehav Rev. 2020;112:519-41.
43. Cudjoe TKM, Selvakumar S, Chung SE, et al. Getting under the skin: Social isolation and biological markers in the National Health and Aging Trends Study. J Am Geriatr Soc. 2021;70:408-14.
44. Van Bogart K, Engeland CG, Sliwinski MJ, et al. The association between loneliness and inflammation: findings from an older adult sample. Front Behav Neurosci. 2022;15:801746.
45. Matthews T, Rasmussen LJH, Ambler A, et al. Social isolation, loneliness, and inflammation: a multi-cohort investigation in early and mid-adulthood. Brain Behav Immun. 2024;115:727-36.
46. Hayek SS, Tahhan AS, Ko Y, et al. Soluble urokinase plasminogen activator receptor levels and outcomes in patients with heart failure. J Card Fail. 2023;29:158-67.
47. Hale S, Yancy CW. New biomarkers in heart failure: the bar is high. J Card Fail. 2023;29:168-70.
48. Koganebuchi K, Kimura R. Biomedical and genetic characteristics of the Ryukyuans: demographic history, diseases and physical and physiological traits. Ann Hum Biol. 2019;46:354-66.
49. Koganebuchi K, Matsunami M, Imamura M, et al. Demographic history of Ryukyu islanders at the southern part of the Japanese Archipelago inferred from whole-genome resequencing data. J Hum Genet. 2023;68:759-67.
50. Cockerham WC, Yamori Y. Okinawa: an exception to the social gradient of life expectancy in Japan. Asia Pac J Clin Nutr. 2008;10:154-8.
51. Sho H. History and characteristics of Okinawan longevity food. Asia Pac J Clin Nutr. 2008;10:159-64.
52. Marinho G, Holdt S, Jacobsen C, Angelidaki I. Lipids and composition of fatty acids of saccharina latissima cultivated year-round in integrated multi-trophic aquaculture. Mar Drugs. 2015;13:4357-74.
53. Sasa M, Inoue I, Shinoda Y, et al. Activating effect of momordin, extract of bitter melon (Momordica Charantia L.), on the promoter of human PPARδ. J Atheroscler Thromb. 2009;16:888-92.
54. Pes GM, Tolu F, Dore MP, et al. Male longevity in Sardinia, a review of historical sources supporting a causal link with dietary factors. Eur J Clin Nutr. 2014;69:411-8.
55. Maioli M, Pes GM, Sanna M, et al. Sourdough-leavened bread improves postprandial glucose and insulin plasma levels in subjects with impaired glucose tolerance. Acta Diabetol. 2008;45:91-6.
56. Fastame MC. Well-being, food habits, and lifestyle for longevity. Preliminary evidence from the sardinian centenarians and long-lived people of the Blue Zone. Psychol Health Med. 2022;27:728-33.
57. Sidore C, Busonero F, Maschio A, et al. Genome sequencing elucidates Sardinian genetic architecture and augments association analyses for lipid and blood inflammatory markers. Nat Genet. 2015;47:1272-81.
58. Poulain M, Herm A. Blue zone, a demographic concept and beyond. Am J Lifestyle Med. 2025:15598276251342502.
59. Poulain M, Herm A, Pes G. The blue zones: areas of exceptional longevity around the world. Vienna Yearb Popul Res. 2013;2013:87-108.
61. Wightman EL, Jackson PA, Khan J, et al. The acute and chronic cognitive and cerebral blood flow effects of a sideritis scardica (greek mountain tea) extract: a double blind, randomized, placebo controlled, parallel groups study in healthy humans. Nutrients. 2018;10:955.
62. Foscolou A, Polychronopoulos E, Paka E, et al. Lifestyle and health determinants of cardiovascular disease among Greek older adults living in Eastern Aegean Islands: an adventure within the MEDIS study. Hellenic J Cardiol. 2016;57:407-14.
63. Chrysohoou C, Pitsavos C, Lazaros G, Skoumas J, Tousoulis D, Stefanadis C. Determinants of all-cause mortality and incidence of cardiovascular disease (2009 to 2013) in older adults: the Ikaria study of the blue zones. Angiology. 2015;67:541-8.
64. Pan C, Cao N. Patterns of social isolation and low social support and frailty trajectories among Chinese older adults. J Appl Gerontol. 2023;42:2325-34.
65. Zhang W, Zhang J, Gao N. Social isolation and cognitive decline in older adults: a longitudinal study across 24 countries. BMC Geriatr. 2025;25:775.
66. Legrand R, Manckoundia P, Nuemi G, Poulain M. Assessment of the health status of the oldest olds living on the Greek Island of Ikaria: a population based-study in a blue zone. Curr Gerontol Geriatr Res. 2019;2019:1-8.
67. Spaulding EM, Astavans A, Bagga A, et al. Sleep and risk for heart disease and stroke. Arterioscler Thromb Vasc Biol. 2016;46:e322869.
68. You S, Zhang HF, Zhang SL, et al. Sleep patterns and traditional cardiovascular health metrics: joint impact on major adverse cardiovascular events in a prospective cohort study. J Am Heart Assoc. 2024;13:e033043.
69. Xie J, Li Y, Zhang Y, et al. Sleep duration and metabolic syndrome: an updated systematic review and meta-analysis. Sleep Med Rev. 2021;59:101451.
70. Mattei J, Hu FB, Campos H. A higher ratio of beans to white rice is associated with lower cardiometabolic risk factors in Costa Rican adults. Am J Clin Nutr. 2011;94:869-72.
71. Yu S, Baylin A, Ruiz-Narváez EA. Micro- and macronutrient intake in elderly costa ricans: the costa rican longevity and healthy aging study (CRELES). Nutrients. 2023;15:1446.
72. Rosero-Bixby L, Dow WH, Rehkopf DH. The nicoya region of costa rica: a high longevity island for elderly males. Vienna Yearb Popul Res. 2014;11:109-36.
73. Gómez Salas G, Arriola Aguirre R, Vindas Meza L, et al. Dietary diversity, anthropometric status and biochemical profile of centenarians of the Nicoya Peninsula, Costa Rica, and their offspring. Rev Esp Geriatr Gerontol. 2024;59:101513.
74. Madrigal-Leer F, Martìnez-Montandòn A, Solìs-Umaña M, et al. Clinical, functional, mental and social profile of the Nicoya Peninsula centenarians, Costa Rica, 2017. Aging Clin Exp Res. 2019;32:313-21.
75. Fraser GE, Shavlik DJ. Ten years of life: is it a matter of choice? Arch Intern Med. 2001;161:1645.
76. Mishra B. Secret of eternal youth; teaching from the centenarian hot spots ("Blue Zones"). Indian J Community Med. 2009;34:273.
77. Rahelić V, Perković T, Romić L, et al. The role of behavioral factors on chronic diseases - practice and knowledge gaps. Healthcare. 2024;12:2520.
78. Santos HD, Alabadi-Bierman A, Paalani M, et al. Living longer and lifestyle: a report on the oldest of the old in the Adventist Health Study-2. J Aging Res Lifestyle. 2025;14:100010.
79. Fraser G. Determinants of ischemic heart disease in Seventh-day Adventists: a review. Am J Clin Nutr. 1988;48:833-6.
80. Orlich MJ, Fraser GE. Vegetarian diets in the Adventist Health Study 2: a review of initial published findings. Am J Clin Nutr. 2014;100:353S-8S.
81. Poulain M, Herm A, Errigo A, et al. Specific features of the oldest old from the Longevity Blue Zones in Ikaria and Sardinia. Mech Ageing Dev. 2021;198:111543.
82. Buettner D. The Blue Zones: 9 lessons for living longer from the people who've lived the longest. Second ed.; 312p. Available from: https://books.google.com/books?hl=en&lr=&id=amJLDXzwpUAC&oi=fnd&pg=PR9&ots=k_Y-ihmGoG&sig=-rpFAPpIG3L-phbYVutMNctHyfk#v=onepage&q&f=false [Last accessed on 15 Jul 2026].
83. Shannon OM, Mendes I, KÖchl C, et al. Mediterranean diet increases endothelial function in adults: a systematic review and meta-analysis of randomized controlled trials. J Nutr. 2020;150:1151-9.
84. Davinelli S, Medoro A, Hu FB, Scapagnini G. Dietary polyphenols as geroprotective compounds: from blue zones to hallmarks of ageing. Ageing Res Rev. 2025;108:102733.
85. Xia M, Zhong Y, Peng Y, Qian C. Olive oil consumption and risk of cardiovascular disease and all-cause mortality: A meta-analysis of prospective cohort studies. Front Nutr. 2022;9:1041203.
86. Nieddu A, Vindas L, Errigo A, Vindas J, Pes GM, Dore MP. Dietary habits, anthropometric features and daily performance in two independent long-lived populations from Nicoya peninsula (Costa Rica) and Ogliastra (Sardinia). Nutrients. 2020;12:1621.
87. Theodoridis X, Chourdakis M, Papaemmanouil A, et al. The effect of diet on vascular aging: a narrative review of the available literature. Life. 2024;14:267.
88. Ataran A, Pompian A, Hajirezaei H, Lodhi R, Javaheri A. Fueling the heart: what are the optimal dietary strategies in heart failure? Nutrients. 2024;16:3157.
89. Lupoli R, Calcaterra I, Ambrosino P, et al. Effects of mediterranean diet on endothelial reactivity in individuals with high cardiometabolic risk: a randomized controlled parallel-group preliminary trial. Biomedicines. 2024;12:2595.
90. Fatima K, Rashid AM, Memon UAA, et al. Mediterranean diet and its effect on endothelial function: a meta-analysis and systematic review. Ir J Med Sci. 2022;192:105-13.
91. Yubero-Serrano EM, Fernandez-Gandara C, Garcia-Rios A, et al. Mediterranean diet and endothelial function in patients with coronary heart disease: an analysis of the CORDIOPREV randomized controlled trial. PLoS Med. 2020;17:e1003282.
92. Hsu C, Huang P, Chiang C, et al. Increased circulating endothelial apoptotic microparticle to endothelial progenitor cell ratio is associated with subsequent decline in glomerular filtration rate in hypertensive patients. PLoS ONE. 2013;8:e68644.
93. Daidone M, Casuccio A, Puleo MG, et al. Mediterranean diet effects on vascular health and serum levels of adipokines and ceramides. PLoS ONE. 2024;19:e0300844.
94. Khandouzi N, Zahedmehr A, Asadian S, Nasrollahzadeh J. Effects of olive oil and flaxseed consumption in a healthy diet on endothelial function, plasma lipids and inflammatory factors of patients with coronary heart disease: a randomized clinical trial. Coron Artery Dis. 2023;34:332-40.
95. Fogacci F, Giovannini M, Di Micoli V, et al. Evaluation of the effect of a dietary supplementation with a red yeast rice and fish oil-containing nutraceutical on lipid pattern, high sensitivity C-reactive protein, and endothelial function in moderately hypercholesterolaemic subjects: a double-blind, placebo-controlled, randomized clinical trial. Arch Med Sci Atheroscler Dis. 2024;8:182-9.
96. Hsu CY, Alzahrani AA, Maabreh HG, et al. Effect of walnut consumption on markers of endothelial function in adults: A systematic review and meta-analysis of randomized controlled trials. Phytother Res. 2024;38:1358-66.
97. Wang Y, Xu D. Effects of aerobic exercise on lipids and lipoproteins. Lipids Health Dis. 2017;16:132.
98. Tucker WJ, Fegers-Wustrow I, Halle M, Haykowsky MJ, Chung EH, Kovacic JC. Exercise for primary and secondary prevention of cardiovascular disease: JACC focus seminar 1/4. J Am Coll Cardiol. 2022;80:1091-106.
99. Ainsworth BE, Haskell WL, Herrmann SD, et al. 2011 compendium of physical activities: a second update of codes and MET values. Med Sci Sports Exerc. 2011;43:1575-81.
100. Shiroma EJ, Lee I. Physical activity and cardiovascular health: lessons learned from epidemiological studies across age, gender, and race/ethnicity. Circulation. 2010;122:743-52.
101. Hellsten Y, Nyberg M. Cardiovascular adaptations to exercise training. Compr Physiol. 2015;6:1-32.
102. Cornelissen VA, Fagard RH. Effects of endurance training on blood pressure, blood pressure–regulating mechanisms, and cardiovascular risk factors. Hypertension. 2005;46:667-75.
103. Martínez-Vizcaíno V, Amaro-Gahete FJ, Fernández-Rodríguez R, Garrido-Miguel M, Cavero-Redondo I, Pozuelo-Carrascosa DP. Effectiveness of fixed-dose combination therapy (polypill) versus exercise to improve the blood-lipid profile: a network meta-analysis. Sports Med. 2021;52:1161-73.
104. Valenzuela PL, Ruilope LM, Santos-Lozano A, et al. Exercise benefits in cardiovascular diseases: from mechanisms to clinical implementation. Eur Heart J. 2023;44:1874-89.
105. Kirwan JP, Sacks J, Nieuwoudt S. The essential role of exercise in the management of type 2 diabetes. Clevel Clin J Med. 2017;84:S15-21.
106. Miura S, Kawanaka K, Kai Y, et al. An increase in murine skeletal muscle peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) mRNA in response to exercise is mediated by β-adrenergic receptor activation. Endocrinology. 2007;148:3441-8.
107. Atherton PJ, Babraj JA, Smith K, Singh J, Rennie MJ, Wackerhage H. Selective activation of AMPK-PGC-1α or PKB-TSC2-mTOR signaling can explain specific adaptive responses to endurance or resistance training-like electrical muscle stimulation. FASEB J. 2005;19:1-23.
108. Tunstall RJ, Mehan KA, Wadley GD, et al. Exercise training increases lipid metabolism gene expression in human skeletal muscle. Am J Physiol Endocrinol Metab. 2002;283:E66-72.
109. Vargas-Ortiz K, Perez-Vazquez V, Diaz-Cisneros FJ, et al. Aerobic training increases expression levels of SIRT3 and PGC-1α in Skeletal muscle of overweight adolescents without change in caloric intake. Pediatr Exerc Sci. 2015;27:177-84.
110. Halling JF, Jessen H, Nøhr-Meldgaard J, et al. PGC-1α regulates mitochondrial properties beyond biogenesis with aging and exercise training. Am J Physiol Endocrinol Metab. 2019;317:E513-25.
111. Cartoni R, Léger B, Hock MB, et al. Mitofusins 1/2 and ERRα expression are increased in human skeletal muscle after physical exercise. J Physiol. 2005;567:349-58.
112. He W, Wang P, Chen Q, Li C. Exercise enhances mitochondrial fission and mitophagy to improve myopathy following critical limb ischemia in elderly mice via the PGC1a/FNDC5/irisin pathway. Skeletal Muscle. 2020;10:25.
113. Axelrod CL, Fealy CE, Mulya A, Kirwan JP. Exercise training remodels human skeletal muscle mitochondrial fission and fusion machinery towards a pro-elongation phenotype. Acta Physiol. 2018;225:e13216.
114. Ristow M, Zarse K, Oberbach A, et al. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc Natl Acad Sci USA. 2009;106:8665-70.
115. Rush JWE, Turk JR, Laughlin MH. Exercise training regulates SOD-1 and oxidative stress in porcine aortic endothelium. Am J Physiol Heart Circ Physiol. 2003;284:H1378-87.
116. Frodermann V, Rohde D, Courties G, et al. Exercise reduces inflammatory cell production and cardiovascular inflammation via instruction of hematopoietic progenitor cells. Nat Med. 2019;25:1761-71.
117. Haykowsky M, Scott J, Esch B, et al. A meta-analysis of the effects of exercise training on left ventricular remodeling following myocardial infarction: start early and go longer for greatest exercise benefits on remodeling. Trials. 2011;12:92.
118. Giallauria F, Lorenzo AD, Pilerci F, et al. Reduction of N terminal-pro-brain (B-type) natriuretic peptide levels with exercise-based cardiac rehabilitation in patients with left ventricular dysfunction after myocardial infarction. Eur J Cardiovasc Prev Rehabil. 2016;13:625-32.
119. Sanchis-Gomar F, Perez-Quilis C, Lucia A. Overexpressing FSTL1 for heart repair. Trends Mol Med. 2016;22:353-4.
120. Christensen RH, Lehrskov LL, Wedell-Neergaard A, et al. Aerobic exercise induces cardiac fat loss and alters cardiac muscle mass through an interleukin-6 receptor-dependent mechanism: cardiac analysis of a double-blind randomized controlled clinical trial in abdominally obese humans. Circulation. 2019;140:1684-6.
121. Recchioni R, Marcheselli F, Antonicelli R, et al. Epigenetic effects of physical activity in elderly patients with cardiovascular disease. Exp Gerontol. 2017;100:17-27.
122. Santos JM, Tewari S, Benite-Ribeiro SA. The effect of exercise on epigenetic modifications of PGC1: the impact on type 2 diabetes. Med Hypotheses. 2014;82:748-53.
123. Herbert C, House M, Dietzman R, Climstein M, Furness J, Kemp-Smith K. Blue zones: centenarian modes of physical activity: a scoping review. Population Ageing. 2022;18:5-41.
124. Fastame MC, Hitchcott PK, Penna MP. The impact of leisure on mental health of Sardinian elderly from the ‘blue zone’: evidence for ageing well. Aging Clin Exp Res. 2017;30:169-80.
125. Willcox DC, Willcox BJ, Sokolovsky J, Sakihara S. The cultural context of “successful aging” among older women weavers in a northern okinawan village: the role of productive activity. J Cross Cult Gerontol. 2007;22:137-65.
126. Pes GM, Errigo A, Tedde P, Dore MP. Sociodemographic, clinical and functional profile of nonagenarians from two areas of sardinia characterized by distinct longevity levels. Rejuvenation Res. 2020;23:341-8.
127. Pes GM, Dore MP, Errigo A, Poulain M. Analysis of physical activity among free-living nonagenarians from a sardinian longevous population. J Aging Phys Act. 2018;26:254-8.
128. Walczak I, Tarnawska M, Stawarska K, et al. Endothelial cell senescence and mitochondrial dysfunction in vascular ageing. Ageing Res Rev. 2026;118:103119.
129. Capurso C, Bellanti F, Lo Buglio A, Vendemiale G. The mediterranean diet slows down the progression of aging and helps to prevent the onset of frailty: a narrative review. Nutrients. 2020;12:35.
130. Ungvari Z, Kaley G, De Cabo R, Sonntag WE, Csiszar A. Mechanisms of vascular aging: new perspectives. J Gerontol A Biol Sci Med Sci. 2010;65A:1028-41.
131. Madamanchi NR, Runge MS. Mitochondrial dysfunction in atherosclerosis. Circ Res. 2007;100:460-73.
132. Kirkman DL, Robinson AT, Rossman MJ, Seals DR, Edwards DG. Mitochondrial contributions to vascular endothelial dysfunction, arterial stiffness, and cardiovascular diseases. Am J Physiol Heart Circ Physiol. 2021;320:H2080-100.
133. Kluge MA, Fetterman JL, Vita JA. Mitochondria and endothelial function. Circ Res. 2013;112:1171-88.
134. Qu K, Yan F, Qin X, et al. Mitochondrial dysfunction in vascular endothelial cells and its role in atherosclerosis. Front Physiol. 2022;13:1084604.
135. Campagna R, Mazzanti L, Pompei V, Alia S, Vignini A, Emanuelli M. The multifaceted role of endothelial Sirt1 in vascular aging: an update. Cells. 2024;13:1469.
136. Mengozzi A, Costantino S, Paneni F, et al. Targeting SIRT1 rescues age- and obesity-induced microvascular dysfunction in ex vivo human vessels. Circ Res. 2022;131:476-91.
137. Kyriazis I, Vassi E, Alvanou M, et al. The impact of diet upon mitochondrial physiology (Review). Int J Mol Med. 2022;50:135.
138. Rossman MJ, Gioscia-Ryan RA, Clayton ZS, Murphy MP, Seals DR. Targeting mitochondrial fitness as a strategy for healthy vascular aging. Clin Sci. 2020;134:1491-519.
139. Mengozzi A, Costantino S, Paneni F, et al. Swimming exercise alleviates endothelial mitochondrial fragmentation via inhibiting dynamin-related protein-1 to improve vascular function in hypertension. Hypertension. 2022;79:e116-28.
140. Rossman MJ, Gioscia-Ryan RA, Santos-Parker JR, et al. Inorganic nitrite supplementation improves endothelial function with aging. Hypertension. 2021;77:1212-22.
141. Han Q, Yu Y, Liu X, et al. The role of endothelial cell mitophagy in age-related cardiovascular diseases. Aging Dis. 2025;16:2151.
142. King-Hudson TJ, Pearson AG, Dunstan-Harrison C, et al. Biomarkers of oxidative and mitochondrial stress are associated with accelerated pace of aging at midlife in a birth cohort. J Gerontol A Biol Sci Med Sci. 2025;80:glaf105.
143. Amorim JA, Coppotelli G, Rolo AP, Palmeira CM, Ross JM, Sinclair DA. Mitochondrial and metabolic dysfunction in ageing and age-related diseases. Nat Rev Endocrinol. 2022;18:243-58.
144. Hubens W, Vallbona-Garcia A, De Coo I, et al. Blood biomarkers for assessment of mitochondrial dysfunction: An expert review. Mitochondrion. 2022;62:187-204.
145. Sanchez-Roman I, Ferrando B, Holst CM, et al. Molecular markers of DNA repair and brain metabolism correlate with cognition in centenarians. GeroScience. 2021;44:103-25.
146. He Q, Wang Y, Zhao F, Wei S, Li X, Zeng G. APE1: a critical focus in neurodegenerative conditions. Biomed Pharmacother. 2024;179:117332.
147. Afshin A, Micha R, Khatibzadeh S, Mozaffarian D. Consumption of nuts and legumes and risk of incident ischemic heart disease, stroke, and diabetes: a systematic review and meta-analysis. Am J Clin Nutr. 2014;100:278-88.
148. Tharrey M, Mariotti F, Mashchak A, Barbillon P, Delattre M, Fraser GE. Patterns of plant and animal protein intake are strongly associated with cardiovascular mortality: the Adventist Health Study-2 cohort. Int J Epidemiol. 2018;47:1603-12.
149. Grosso G, Godos J, Lamuela-Raventos R, et al. A comprehensive meta-analysis on dietary flavonoid and lignan intake and cancer risk: level of evidence and limitations. Mol Nutr Food Res. 2017;61:1600930.
150. Kavyani Z, Musazadeh V, Fathi S, Hossein Faghfouri A, Dehghan P, Sarmadi B. Efficacy of the omega-3 fatty acids supplementation on inflammatory biomarkers: an umbrella meta-analysis. Int Immunopharmacol. 2022;111:109104.
151. Hornero-Ramirez H, Aubin A, Michalski M, Vinoy S, Caussy C, Nazare J. Multifunctional dietary interventions, low-grade inflammation and cardiometabolic profile: a scoping review. Front Immunol. 2024;15:1304686.
152. Di Giosia P, Stamerra CA, Giorgini P, Jamialahamdi T, Butler AE, Sahebkar A. The role of nutrition in inflammaging. Ageing Res Rev. 2022;77:101596.
153. Mchill AW, Butler MP. Eating around the clock: circadian rhythms of eating and metabolism. Annu Rev Nutr. 2024;44:25-50.
154. Davis R, Rogers M, Coates AM, Leung GKW, Bonham MP. The impact of meal timing on risk of weight gain and development of obesity: a review of the current evidence and opportunities for dietary intervention. Curr Diab Rep. 2022;22:147-55.
155. Grimaldi D, Reid KJ, Abbott SM, Knutson KL, Zee PC. Sleep-aligned extended overnight fasting improves nighttime and daytime cardiometabolic function. Arterioscler Thromb Vasc Biol. 2026;46:e323355.
156. Frias-Toral E, Reytor-González C, Annunziata G, et al. Nutrition and longevity - diet in centenarians. J Transl Med. 2026;24:299.
157. Guan G, Chen Y, Dong Y. Unraveling the AMPK-SIRT1-FOXO pathway: the in-depth analysis and breakthrough prospects of oxidative stress-induced diseases. Antioxidants. 2025;14:70.
158. Trefts E, Shaw RJ. AMPK: restoring metabolic homeostasis over space and time. Mol Cell. 2021;81:3677-90.
159. Liu GY, Sabatini DM. mTOR at the nexus of nutrition, growth, ageing and disease. Nat Rev Mol Cell Biol. 2020;21:183-203.
160. Sun C, Bai S, Liang Y, et al. The role of Sirtuin 1 and its activators in age-related lung disease. Biomed Pharmacother. 2023;162:114573.
161. Barcena ML, Aslam M, Norman K, Ott C, Ladilov Y. Role of AMPK and sirtuins in aging heart: basic and translational aspects. Aging Dis. 2024;16:3335-60.
163. Hou Y, Hu S, Li X, He W, Wu G. Amino acid metabolism in the liver: nutritional and physiological significance. Adv Exp Med Biol. 2020;1265:21-37.
164. Matsumoto T, Nakamura K, Matsumoto H, et al. Bolus ingestion of individual branched-chain amino acids alters plasma amino acid profiles in young healthy men. SpringerPlus. 2014;3:35.
165. Jenkins DJA, Kendall CWC, Augustin LSA, et al. Effect of legumes as part of a low glycemic index diet on glycemic control and cardiovascular risk factors in type 2 diabetes mellitus: a randomized controlled trial. Arch Intern Med. 2012;172:1653.
166. Viguiliouk E, Stewart SE, Jayalath VH, et al. Effect of replacing animal protein with plant protein on glycemic control in diabetes: a systematic review and meta-analysis of randomized controlled trials. Nutrients. 2015;7:9804-24.
167. Mudryj AN, Yu N, Aukema HM. Nutritional and health benefits of pulses. Appl Physiol Nutr Metab. 2014;39:1197-204.
168. Anderson JW, Major AW. Pulses and lipaemia, short- and long-term effect: Potential in the prevention of cardiovascular disease. Br J Nutr. 2007;88:263-71.
169. Lu T, Xie Y, Wang Y, et al. Protein restriction reprograms the multi-organ proteomic landscape of mouse aging. Cell. 2025;188:7309-26.e20.
170. Jackman SR, Witard OC, Philp A, Wallis GA, Baar K, Tipton KD. Branched-chain amino acid ingestion stimulates muscle myofibrillar protein synthesis following resistance exercise in humans. Front Physiol. 2017;8:390.
171. Huang J, Kwok AJ, Li JCY, et al. Body-wide multi-omic counteraction of aging with GLP-1R agonism. Cell Metab. 2025;37:2362-80.e8.
172. Brandhorst S, Longo VD. Protein quantity and source, fasting-mimicking diets, and longevity. Adv Nutr. 2019;10:S340-50.
174. Wali JA, Solon-Biet SM, Freire T, Brandon AE. Macronutrient determinants of obesity, insulin resistance and metabolic health. Biology. 2021;10:336.
175. De Chiara F, Ureta Checcllo C, Ramón Azcón J. High protein diet and metabolic plasticity in non-alcoholic fatty liver disease: myths and truths. Nutrients. 2019;11:2985.
176. Ekmekcioglu C. Nutrition and longevity - from mechanisms to uncertainties. Crit Rev Food Sci Nutr. 2019;60:3063-82.
177. Polom J, Boccardi V. Employing nutrition to delay aging: a plant-based telomere-friendly dietary revolution. Nutrients. 2025;17:2004.
178. Yaskolka Meir A, Keller M, Hoffmann A, et al. The effect of polyphenols on DNA methylation-assessed biological age attenuation: the DIRECT PLUS randomized controlled trial. BMC Med. 2023;21:364.
179. Maegawa S, Lu Y, Tahara T, et al. Caloric restriction delays age-related methylation drift. Nat Commun. 2017;8:539.
180. Qiu Z, Huang EYZ, Li Y, et al. Beneficial effects of time-restricted fasting on cardiovascular disease risk factors: a meta-analysis. BMC Cardiovasc Disord. 2024;24:210.
181. Bensalem J, Teong XT, Hattersley KJ, et al. Intermittent time-restricted eating may increase autophagic flux in humans: an exploratory analysis. J Physiol. 2025;603:3019-32.
182. Xinyan C, Yajie W, Shangfan H, et al. mTOR-autophagy axis regulation by intermittent fasting promotes skeletal muscle growth and differentiation. Nutr Metab. 2025;22:109.
183. Ham DJ, Börsch A, Chojnowska K, et al. Distinct and additive effects of calorie restriction and rapamycin in aging skeletal muscle. Nat Commun. 2022;13:2025.
184. Dai D, Kang P, Bai H. The mTOR signaling pathway in cardiac aging. J Cardiovasc Aging. 2023;3:24.
185. Ataman M, Mittal N, Tintignac L, et al. Calorie restriction and rapamycin distinctly mitigate aging-associated protein phosphorylation changes in mouse muscles. Commun Biol. 2024;7:974.
186. Demontis F, Perrimon N. FOXO/4E-BP signaling in drosophila muscles regulates organism-wide proteostasis during aging. Cell. 2010;143:813-25.
187. Longo VD, Di Tano M, Mattson MP, Guidi N. Intermittent and periodic fasting, longevity and disease. Nat Aging. 2021;1:47-59.
188. Espinoza SE, Park S, Connolly G, et al. Effect of fasting-mimicking diet on markers of autophagy and metabolic health in human subjects. GeroScience. 2025:2035.
189. Popa AD, Gherasim A, Mihalache L, Arhire LI, Graur M, Niță O. Fasting mimicking diet for metabolic syndrome: a narrative review of human studies. Metabolites. 2025;15:150.
190. Crupi AN, Haase J, Brandhorst S, Longo VD. Periodic and intermittent fasting in diabetes and cardiovascular disease. Curr Diab Rep. 2020;20:83.
191. Burns L, Cooper S, Sarmad S, et al. Effects of fasting-mimicking diets with low and high protein content on cardiometabolic health and autophagy: a randomized, parallel group study. Clin Nutr. 2025;52:299-312.
192. Navarro-Ibarra MJ, Hernández J, Caire-Juvera G. Diet, physical activity and telomere length in adults. Nutr Hosp. 2019;36:1403-17.
193. Li X, Li C, Zhang W, Wang Y, Qian P, Huang H. Inflammation and aging: signaling pathways and intervention therapies. Sig Transduct Target Ther. 2023;8:239.
194. Arai Y, Martin-Ruiz CM, Takayama M, et al. Inflammation, but not telomere length, predicts successful ageing at extreme old age: a longitudinal study of semi-supercentenarians. EBioMedicine. 2015;2:1549-58.
195. Sayed N, Huang Y, Nguyen K, et al. An inflammatory aging clock (iAge) based on deep learning tracks multimorbidity, immunosenescence, frailty and cardiovascular aging. Nat Aging. 2021;1:598-615.
196. Zhou L, Ge M, Zhang Y, et al. Centenarians alleviate inflammaging by changing the ratio and secretory phenotypes of t helper 17 and regulatory T cells. Front Pharmacol. 2022;13:877709.
197. Engelbrecht H, Merrill SM, Gladish N, et al. Sex differences in epigenetic age in Mediterranean high longevity regions. Front Aging. 2022;3:1007098.
198. Sotgia S, Zinellu A, Mangoni AA, et al. Cellular immune activation in Sardinian middle-aged, older adults and centenarians. Exp Gerontol. 2017;99:133-7.
199. Gonda K, Kanazawa H, Maeda G, et al. Ingestion of okinawa island vegetables increases IgA levels and prevents the spread of influenza RNA viruses. Nutrients. 2021;13:1773.
200. Tomori M, Nagamine T, Miyamoto T, Iha M. Effects of ingesting fucoidan derived from cladosiphon okamuranus tokida on human NK cells: a randomized, double-blind, parallel-group, placebo-controlled pilot study. Mar Drugs. 2021;19:340.
201. Mcewen LM, Morin AM, Edgar RD, et al. Differential DNA methylation and lymphocyte proportions in a Costa Rican high longevity region. Epigenet Chromatin. 2017;10:21.
202. Pinti M, Gibellini L, Lo Tartaro D, et al. A comprehensive analysis of cytokine network in centenarians. Int J Mol Sci. 2023;24:2719.
203. Zhao Y, Liu Y. Longevity factor FOXO3: a key regulator in aging-related vascular diseases. Front Cardiovasc Med. 2021;8:778674.
204. Torigoe TH, Willcox DC, Shimabukuro M, et al. Novel protective effect of the FOXO3 longevity genotype on mechanisms of cellular aging in Okinawans. NPJ Aging. 2024;10:18.
205. White RR, Maslov AY, Lee M, Wilner SE, Levy M, Vijg J. FOXO3a acts to suppress DNA double-strand break-induced mutations. Aging Cell. 2020;19:e13184.
206. Inci G, Warkad MS, Kang B, et al. FOXO3a upregulates DNA repair activities by transcriptional activation of target genes and provides the resistance to gamma radiation and the extension of lifespan in mouse. Biogerontology. 2025;26:196.
207. Toribio-Fernández R, Tristão-Pereira C, Carlos Silla-Castro J, et al. Apolipoprotein E-ε2 and resistance to atherosclerosis in midlife: the PESA observational study. Circ Res. 2024;134:411-24.
208. Kim H, Devanand DP, Carlson S, Goldberg TE. Apolipoprotein E genotype e2: neuroprotection and its limits. Front Aging Neurosci. 2022;14:919712.
209. Melis M, Errigo A, Crnjar R, Pes GM, Tomassini barbarossa I. TAS2R38 bitter taste receptor and attainment of exceptional longevity. Sci Rep. 2019;9:18047.
210. Paika S, Machini M, Parmar MS. Molecular mechanisms of drug-induced hemolysis in G6PD deficiency: mechanistic insights. Oxid Med Cell Longev. 2025;2025:7041213.
211. Mak GK, Shah M. Glucose-6-phosphate dehydrogenase deficiency; 2025. Available from: https://www.ncbi.nlm.nih.gov/books/NBK470315/ [Last accessed on 15 Jul 2026].
212. Austad SN, Pes GM. The validity of Blue Zones demography: a response to critiques. Gerontologist. 2025;65:gnaf246.
214. Errigo A, Dore MP, Mocci G, Pes GM. Lack of association between common polymorphisms associated with successful aging and longevity in the population of Sardinian Blue Zone. Sci Rep. 2024;14:30773.
215. Candal-Pedreira C, Rey-Brandariz J, Martín-Gisbert L, et al. Blue zones, an analysis of existing evidence through a scoping review. Aging Dis. 2025;17:1335-46.
216. Helte E, Säve-Söderbergh M, Larsson SC, Åkesson A. Calcium and magnesium in drinking water and risk of myocardial infarction and stroke—a population-based cohort study. Am J Clin Nutr. 2022;116:1091-100.
217. Aliberti SM, Funk RHW, Ciaglia E, et al. Old, nonagenarians, and centenarians in cilento, Italy and the association of lifespan with the level of some physicochemical elements in tap drinking water. Nutrients. 2023;15:218.
218. Aliberti SM, Donato A, Funk RHW, Capunzo M. A narrative review exploring the similarities between cilento and the already defined “blue zones” in terms of environment, nutrition, and lifestyle: can cilento be considered an undefined “blue zone”? Nutrients. 2024;16:729.
219. Hori M, Shozugawa K, Sugimori K, Watanabe Y. A survey of monitoring tap water hardness in Japan and its distribution patterns. Sci Rep. 2021;11:13546.
220. Pietri P, Papaioannou T, Stefanadis C. An old clue to the secret of longevity. Nature. 2017;544:416.
221. Newman SJ. Supercentenarian and remarkable age records exhibit patterns indicative of clerical errors and pension fraud. Dev Biol. 2019:704080.
222. Poulain M, Herm A. Exceptional longevity in Okinawa: demographic trends since 1975. J Intern Med. 2024;295:387-99.
223. Hao L, Xu X, Dupre ME, et al. Adequate access to healthcare and added life expectancy among older adults in China. BMC Geriatr. 2020;20:129.
224. Caruso C, Accardi G, Aiello A, Calabrò A, Zarcone R, Candore G. The longevity of blue zones: myth or reality. J Gerontol Geriatr. 2025;73:55-66.
225. Echeverry Raad J, Sturmberg JP. Red zones: the true color behind the myth of blue zones geographic longevity. Rev Publica. 2025;27:1-7.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].