


Circular RNAs as potential therapeutics and novel biomarkers for heart failure
 0
0 
Abstract
Heart failure (HF) is a type of chronic pathological remodeling in which the heart fails to efficiently supply an adequate amount of blood to fulfill the body’s requirements. Circular RNAs (circRNAs) are recognized as specific types of long noncoding RNAs that are involved in the regulation of the pathogenesis of cardiovascular diseases, including cardiac remodeling and HF. In this review, knowledge about the origin, characteristics, and function of circRNAs is summarized. The regulatory function of circRNAs in cardiac remodeling mechanisms, including cell death, cardiac hypertrophy, fibrosis and mitochondrial metabolism, is reviewed. Finally, future directions for investigating the diagnostic and pharmacological potential of circRNAs in HF are proposed.
Keywords
INTRODUCTION
Heart failure (HF) is typically characterized by pathological remodeling with a decrease in cardiac output to fulfill the metabolic demands of the body and is often promoted by diseases that contribute to pressure or volume overload, such as hypertension, myocardial infarction (MI), valvular heart disease and
Cardiac remodeling initially protects the heart as an adaptive response, which refers to a series of compensatory changes in the size, shape, and function of the myocardium in response to a variety of stimuli[3], but when these changes are prolonged, progressive cardiac dysfunction occurs. MI, ischemia/reperfusion (I/R), injury following coronary revascularization and on-pump cardiac surgery, pressure and volume overload, and neuroendocrine activation are high-risk stimuli that trigger cardiac remodeling[4]. Histopathological hallmarks, including cell death, defective autophagy, cardiac hypertrophy, fibrosis, inflammation and metabolic and mitochondrial dysfunction, are involved in cardiac remodeling[5].
RNA-sequencing (RNA-Seq) technology has been applied to identify novel transcripts, such as noncoding RNAs (ncRNAs) and alternative splicing isoforms of each transcript[6]. Accumulating evidence has revealed that ncRNAs, such as microRNAs (miRNAs), PIWI-interacting RNAs (piRNAs), circular RNAs (circRNAs), and long noncoding RNAs (lncRNAs), are pivotal regulators of gene expression involved in cardiac remodeling[7-9]. CircRNAs, with a covalently closed loop structure, confer greater stability than linear RNAs. Most circRNAs are present at relatively low levels in cells, and some of them represent significant or even predominant transcript isoforms[10]. CircRNAs exert their biological functions through specifically combining miRNAs, binding to RNA-binding proteins (RBPs), translating proteins, and modulating gene transcription[11,12].
The present review focuses on the functional roles of circRNAs in cardiac remodeling leading to HF. CircRNA biogenesis and its mechanisms of action are briefly overviewed. Moreover, a landscape of the newest findings is described with respect to the role of circRNAs in cardiac remodeling and HF, with a focus on their regulation of cardiac cell death, cardiac hypertrophy, fibrosis and mitochondrial metabolism dysregulation. Furthermore, the current knowledge of the potential of circRNAs for diagnostic and therapeutic purposes is reviewed, and directions for future work are proposed to explore the pharmacological potential of circRNAs in HF.
CircRNA BIOGENESIS AND MECHANISMS OF ACTION
CircRNAs are covalently closed RNAs that are resistant to the 3′-exonuclease RNase R[13]. CircRNAs are formed from precursor mRNAs (pre-mRNAs) through the back-splicing. CircRNA molecules are primarily generated via the following two modes of back-splicing: (1) Intron complementary sequence-driven circularization. Reverse complementary repeat sequences (such as Alu elements) exist within the flanking introns. These sequences can form secondary structures such as “stem loops” through base pairing, thereby bringing the splice sites into proximity to initiate circularization. (2) RNA-binding protein (RBP)-driven circularization. Specific RBPs (e.g., QKI and FUS) bind to specific motifs within introns. Through protein dimerization or conformational changes, these RBPs bridge two distal splice sites, inducing circularization.
These two circularization modes can result in three subclasses of circRNAs: intronic circRNAs (IcircRNAs), exonic circRNAs (EcircRNAs), and exon-intron circRNAs (EicircRNAs). IcircRNAs are generated via a lariat-driven mechanism, whereas EcircRNAs and EicircRNAs are produced through intron circularization[14].
An increasing number of studies have revealed different roles of circRNAs, ranging from the regulation of transcription to the sponging of RNA/protein and translation of protein. The biological functions of circRNAs are diverse and primarily involve four major mechanisms of action: (1) microRNA sponges: The “competing endogenous RNA (ceRNA)” hypothesis posits that circRNAs contain binding sites for specific miRNAs. Some cytoplasmic circRNAs contain multiple miRNA binding sites, allowing a single circRNA to bind a number of miRNAs to block the functions of target miRNAs[15]. (2) Protein interactions: The interactions between circRNAs and proteins can be categorized into three scenarios: a circRNA binds to two proteins, thereby cementing their interaction; a circRNA binds to one protein and enhances or disrupts its interaction with another protein; and a circRNA binds to two proteins that interact with each other, thereby abolishing the existing interaction[16]. (3) Translating proteins or peptides: Due to the lack of a 5' cap structure, circRNAs employ cap-independent translation. The internal ribosome entry site (IRES) contained in circRNA can recruit ribosomes to initiate protein translation. Nucleotide sites modified by m6A can be recognized by m6A reader proteins, thereby recruiting translation initiation factors to start translation of the downstream open reading frame (ORF). Some circRNAs with a continuous ORF can undergo multiple rounds of the rolling circle translation along with the circular template by ribosomes[17]. (4) Regulating gene transcription: IcircRNAs or EicircRNAs in the nucleus can interact with U1 small nuclear ribonucleoprotein (U1 snRNP) to form a complex that acts on RNA polymerase II, thereby initiating gene transcription. Alternatively, IcircRNAs or EicircRNAs may interact with RNA Pol II to modulate gene expression[18] [Figure 1].
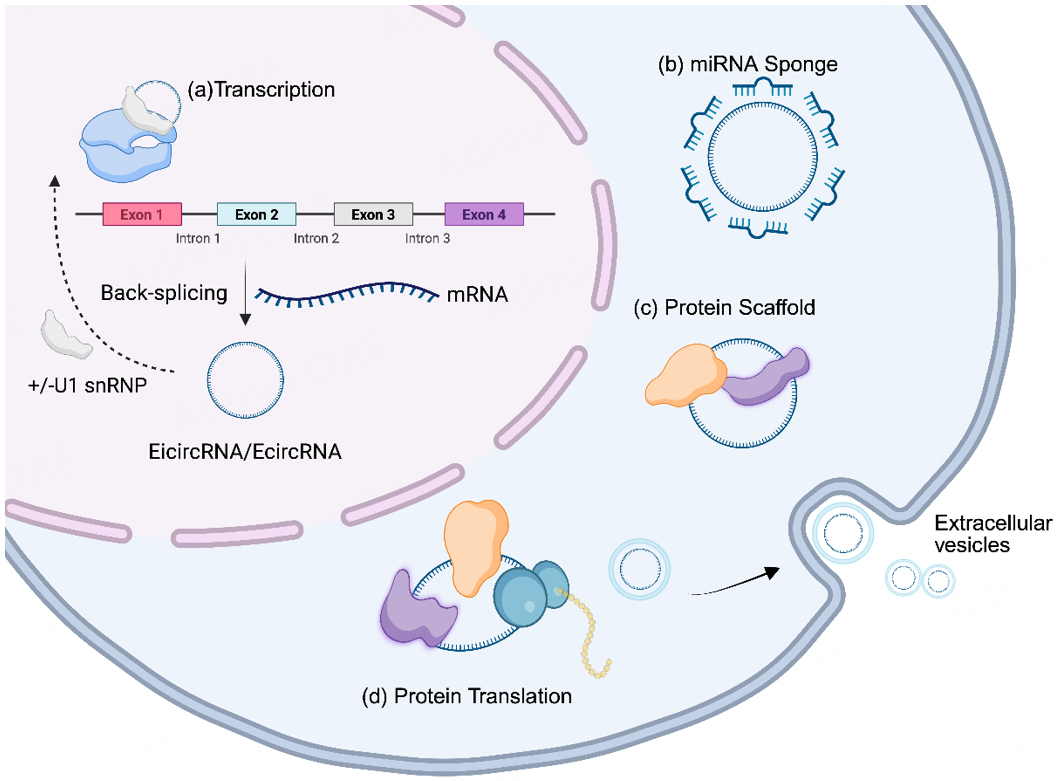
Figure 1. The types of functions of circRNAs in the nucleus and cytoplasm. (a) Modulate Transcription. The EicircRNA/EcircRNA binds U1 snRNP and RNA polymerase II to regulate the parental gene transcription in the nucleus. (b) miRNA sponge. In the cytoplasm, circRNAs sponge the target miRNAs, sequestering and inhibiting miRNA function, thereby regulating gene expression. (c) Protein scaffold. CircRNAs bind to RBPs and serve as scaffolds for the assembly of protein complexes, participating in signaling or cellular structure maintenance. (d) Protein translation. CircRNAs encode functional peptides or proteins via mechanisms such as internal ribosome entry sites (IRES), participating in cellular physiological processes. Created in BioRender, https://BioRender.com/mc4x7y4.
CircRNAs INVOLVED IN CARDIAC REMODELING AND HF
The increasing evidence has shown that circRNAs play important roles in the pathophysiological mechanisms of cardiac remodeling and HF. Hereafter, studies assessing the functional roles of circRNAs in myocardial remodeling and HF using both in vitro and in vivo models are presented [Tables 1 and 2,
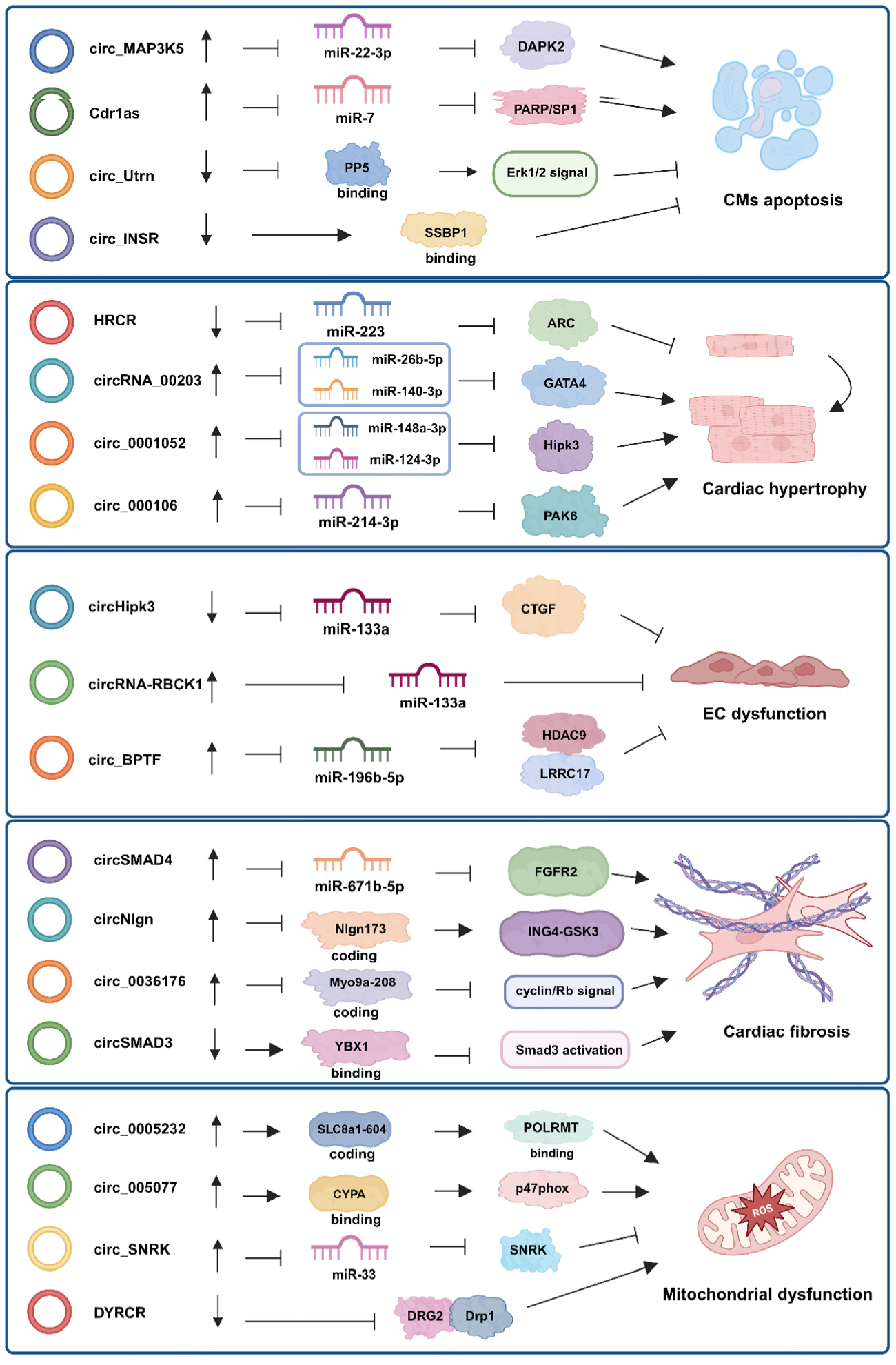
Figure 2. CircRNA regulatory axes in cardiomyocyte apoptosis, cardiac hypertrophy, endothelial cell dysfunction, cardiac fibrosis and mitochondrial dysfunction. CircRNAs stimulate or inhibit mechanisms of pathological cardiac remodeling via sponging microRNAs, binding RNA-binding proteins or translating proteins. Created in BioRender, https://BioRender.com/usskcio.
CircRNAs in cardiomyocyte and endothelial cell biology
| circRNA | Source | Expression change | Function | Down-stream target | Reference |
| Cardiac cell death | |||||
| Cdr1as | CMs, MI mice | Up | Promoting MI-induced apoptosis | miR-7/PARP and SP1 | [20] |
| circ_ZNF609 | I/R mice, DOX-induced cardiotoxicity | Up | Inducing CM apoptosis and promoting ROS production | YTHDF3-FTO | [21,22] |
| circSNRK | MI mice | Down | Reducing apoptosis of CMs | miR-103-3p/SNRK-GSK-3β/β-catenin | [23] |
| DICAR | Diabetic mouse, mouse CMs | Down | Suppressing CM pyroptosis | VCP | [24] |
| circ_0001206 | H9C2 cells | Down | Promoting cell viability | miR-665/CRKL | [25] |
| circ_BNIP3 | Rat CMs | Up | Promoting hypoxia-activated apoptosis | miR-27a-3p/BNIP3 | [26] |
| circ_Fbxl5 | I/R mice, A/R-treated NMVMs and CFs | Up | Inducing CM apoptosis and increasing infarct size | miR-146a/MED1 | [27] |
| circ_Utrn | I/R mice, hESC-CMs | Down | Attenuating I/R-induced apoptosis | PP5 | [28] |
| CNEACR (mmu_circ_000338) | I/R mice | Down | Promoting cell survival | CNEACR/HDAC7/FOXO2/RIPK3 | [29] |
| FEACR | I/R mice | Down | Inhibiting ferroptosis in CMs | NAMPT-Sirt1-FOXO1-FTH1 | [30] |
| circ_INSR | DOX-induced HF | Down | Enhancing cell viability | SSBP1 | [31] |
| circ_MAP3K5 | DCM rats | Up | Promoting CMs apoptosis | miR-22-3p/DAPK2 | [32] |
| Cardiac hypertrophy | |||||
| circCDYL | TAC mice, Ang II-treated rat CMs | Up | Promoting cardiac hypertrophy | tCDYL-100/REST-CDYL-EHMT2/Rhoa-Nppb | [35] |
| circRYR2 (mmu_circ_0000431) | TAC mice, Mouse CMs | Down | Preventing cardiac hypertrophy | SERCA2a to improve Ca2+ handling | [36] |
| circ_0001052 | TAC mice, CMs, HL-1 cells | Up | Promoting cardiac hypertrophy | miR-148a-3p, -124-3p/Hipk3 | [37] |
| HRCR (circ_012559) | ISO-injected mice and TAC mice, mouse CMs | Down | Reducing cardiac hypertrophy | miR-223/ARC | [38] |
| circNfix | Ang II-treated MCs | Down | Preventing cardiac hypertrophy | miR-145-5p/ATF3 | [40] |
| circRNA_000203 | Ang II-infused mice, mouse CMs treated with Ang II | Up | Enhancing myocardial hypertrophy | miR-26b-5p, -140-3p/GATA4 | [41] |
| circ_0001006 | TAC mice, Ang II-treated CMs | Up | Exacerbating cardiac hypertrophy | miR-214-3p/PAK6 | [42] |
| EC dysfunction | |||||
| circHipk3 | MI mice, mouse CMs | Down | Promoting EC proliferation and angiogenesis | miR-133a/CTGF | [45] |
| circRNA-RBCK1 (circRNA_102979) | Statin-induced human ECs, HFpEF mice | Up | Preventing diastolic dysfunction in HFpEF mice | miR-133a | [46] |
| circ_BPTF | LV of ischaemic HF patients; HUVEC cells | Up | Inducing endothelial cell proliferation | miR-196b-5p/HDAC9 and LRRC17 | [49] |
CircRNAs in fibrosis and mitochondrial function
| circRNA | Source | Expression change | Function | Down-stream target | Reference |
| Cardiac fibrosis | |||||
| circ_Yap | Heart samples of HF patients; TAC mice | Down | Attenuating heart fibrosis | TMP4/ACTG | [51] |
| circNAB1 | AF patient specimens | Down | Atrial fibrosis and AF susceptibility | Encoding NAB1-356 | [52] |
| circ_CAMTA1 | Atrial tissue of AF patients, Ang II-treated AFs | Up | Stimulating expression of fibrotic markers | miR-214-3p/TGFβ1R | [53] |
| circNlgn | Cardiac tissue of CHD patients | Up | Inducing cardiac fibrosis and HF | NIgn173-ING4/C8orf44-SGK3 | [54] |
| circSMAD3 | Murine with HF | Down | Mitigating cardiac fibrosis | YBX1/Smad3 | [55] |
| circHIPK2 | Murine MI model | Up | Promoting cardiac fibrosis, inflammation | Interacts with G3BP1 | [56] |
| circSMAD4 | TAC mice | Up | Promoting cardiac fibrosis | miR-671-5p/FGFR2 | [57] |
| circSnap47 | Myocardium of HF patients, H9c2 cells | Up | circSnap47 KD suppressing HF | miR-223-3p/MAPK | [58] |
| circIGF1R (has_circ_0005035) | Cardiac tissue of HF patients | Up | Inhibiting cardiac fibrosis | Via AZGP1 inhibiting glycolysis | [59] |
| circPVT1 | Ischemic HF patients | Up | Promoting cardiac fibrosis | Sponging miR-30a-5p, -125b-5p | [60] |
| circMap4K2 | MI mice | Up | Attenuating residual remodeling | MiR-106a-3p/Azin1 | [61] |
| circ_0002295 | Patients with cardiac fibrosis | Up | Facilitating cardiac fibrosis | miR-1287/CXCR2 | [62] |
| circUbe3a | AMI mice | Up | Exacerbating myocardial fibrosis | miR-138-5p/RhoC | [63] |
| circ-CELF1 | MI mice, TGF-β1-treated CFs | Up | Exacerbating cardiac fibrosis | BRPF3/KAT7-Celf1/Smad7 | [64] |
| circ_0036176 | Myocardium of HF patients | Up | Suppressing CFs proliferation | Myo9a-208/Cyclin/Rb | [65] |
| circ_Helz | AMI/Ang II-induced mice; TGFβ-treated CFs | Up | Promoting cardiac fibrosis | YAP | [66] |
| circ_LAS1L | Blood samples of AMI patients; HCFs | Down | Inhibiting CF proliferation and migration | miR-125b/SFRP5 | [67] |
| Aberrant metabolism /mitochondrial dysfunction | |||||
| DICAR | Diabetic mouse model, mouse CMs | Down | Suppressing CM pyroptosis | VCP | [24] |
| circ_INSR | DOX-induced human heart tissues; DOX-induced mouse model | Down | Attenuating DOX-mediated apoptosis | SSBP1 | [31] |
| circ_0005232 | Cardiac tissue of HF patients | Up | Enhancing cardiac remodeling and HF | SLC8a1-604-POLRMT/gene transcription, ATP synthesis | [72] |
| circ_005077 | HFD-fed rats | Up | Enhancing myocardial lipotoxicity | CYPA-p47phox-NADPH-ROS | [74] |
| PYRCR | I/R mice, H/R MCs | Down | Attenuating MC pyroptosis and myocardial damage | DRG2-Drp1-mitochondrial fission | [73] |
| circ-SNRK | MI mice, CMs | Down | Improving cardiac function | miR-33-SNRK | [75] |
Cardiomyocyte death
It is well known that the pathological progression of cardiac diseases leading to chronic HF is marked by the absence of cardiomyocytes (CMs) or vascular cells. Different types of cell death, including apoptosis, necroptosis, pyroptosis and autophagy, contribute to cardiomyocyte death[5]. Various stressors, including cytokines, increased oxidative stress and DNA damage, promote cell death in cardiomyocytes, and inhibition of cell death is cardioprotective against the development of HF[19]. Different circRNAs have been demonstrated to regulate CM death[20-32].
Cdr1as, which is upregulated in the hearts of MI mice and in hypoxia-treated CMs, induces CM apoptosis in MI by sponging miR-7. Geng et al. reported that overexpression of Cdr1as in a mouse CM cell line increased apoptosis, which could be abolished by miR-7a administration. Specificity protein 1 (SP1) and PARP were validated as relevant target genes of miR-7a[20]. In particular, Cdr1as overexpression in mice exacerbated MI injury and increased the expression of PARP and SP1, suggesting that these proteins are targets of the Cdr1as-miR-7 network[20].
Recently, circ-ZNF609 knockdown in an I/R injury mouse model was shown to reduce left ventricular (LV) dysfunction and promote CM proliferation. Circ-ZNF609 overexpression exacerbated CM apoptosis, whereas knockdown of circ-ZNF609 decreased YTHDF3 expression, promoted the binding of YTHDF1 to Yap mRNA and increased Yap translation. Circ-ZNF609 overexpression interferes with the cross-talk between Hippo-YAP and Akt signaling by suppressing YAP expression[21]. Moreover, knockdown of circ-ZNF609 attenuated doxorubicin-induced CM apoptosis. Mechanistically, the absence of circ-ZNF609 reduced reactive oxygen species (ROS) accumulation and ameliorated mitochondrial iron overload by inhibiting Fto mRNA degradation[22]. These data suggest that circ-ZNF609 is a promising target for the treatment of cardiac I/R injury and doxorubicin-induced cardiotoxicity.
Zhu et al.[23] elucidated the role of circSNRK in the modulation of cardiomyocyte survival. CircSNRK expression was decreased in rats with MI, and overexpression of circSNRK attenuated cardiomyocyte apoptosis. Adeno-associated virus 9 (AAV9)-mediated exogenous overexpression of circSNRK decreased cardiomyocyte apoptosis and promoted proliferation. Mechanistically, circSNRK inhibits cardiomyocyte apoptosis by sponging miR-103-3p to increase the level of SNRK, which increases the phosphorylation of GSK3β.
In another study, Yuan et al. reported a conserved circRNA, DICAR, that has an inhibitory effect on diabetic cardiomyopathy (DCM)[24]. DICAR was downregulated in diabetic mouse hearts, and DICAR overexpression inhibited but DICAR knockdown enhanced diabetic cardiomyocyte pyroptosis. Spontaneous cardiac dysfunction occurred in DICAR-deficient (DICAR+/-) mice but was ameliorated in transgenic Tg-DICAR mice. Mechanistically, DICAR-VCP-Med12 degradation participated in DICAR-mediated effects. Moreover, this study indicated that DICAR, as well as the synthesized DICAR junction component (DICAR-JP), may be potential drug candidates for the treatment of DCM.
Cardiac hypertrophy
Cardiac hypertrophy is the most important pathological basis for myocardial remodeling and HF, cardiovascular morbidity and mortality. Pathological hypertrophy is accompanied by fibrosis, increased proinflammatory cytokines, and cellular impairment, as well as undesirable epigenetic changes that participate in maladaptive cardiac remodeling[33,34].
CircRNAs have been shown to be involved in the modulation of the pathological processes of myocardial hypertrophy[35-42]. Li et al. reported that circCDYL, derived from the CDYL gene, promoted cardiac hypertrophy through the N6-methyladenosine-circRNA-histone methylation axis[35]. Functional assays revealed that circCDYL effectively induced myocardial hypertrophy. CircCDYL encoded a CDYL peptide (tCDYL-100), and this process of translation was promoted by the N6-methylation of circCDYL. tCDYL-100 competed with CDYL to combine with REST to disrupt the formation of the REST-CDYL-EHMT2 complex, leading to the transcriptional activation of rhoa and nppb. Additionally, the absence of circCDYL inhibited Ang II-induced myocardial hypertrophy, whereas exogenous overexpression of tCDYL-100 promoted myocardial hypertrophy[35].
Pan et al. elucidated the role of circRYR2 (mmu_circ_0000431) in the process of myocardial hypertrophy[36]. The expression of the human circRYR2 homolog hsa_circ_0112647 was decreased in failing human hearts. The absence of circRYR2 in cardiomyocytes led to hypertrophic responses, whereas circRYR2 overexpression prevented cytokine-induced cardiomyocyte hypertrophy. Mechanistically, circRYR2 modulated the expression of SERCA2a, and circRYR2 knockdown impaired Ca2+ activity, but circRYR2 overexpression improved Ca2+ activity[36].
Recent research revealed that HIPK3 gene-derived circ_0001052 was upregulated in both TAC-treated mice and Ang II-treated CMs and that its depletion abrogated the Ang II-induced increase in cell surface area. Mechanistically, circ_0001052 stabilized Hipk3 mRNA by sequestering miR-148a-3p/miR-124-3p and binding to the Srsf1 protein, thus promoting CM hypertrophy[37].
Endothelial cell dysfunction
Coronary microcirculation enables the exchange of nutrients and waste metabolites between the vasculature and cardiomyocytes. Pathologic conditions, including hypertension, cardiomyopathy, and MI, enhance the process of microcirculatory remodeling. Microvascular dysfunction typically increases the heterogeneity of blood flow and local hypoxia in the heart, contributing to contractile and diastolic abnormalities[43,44].
Si et al. investigated the effect of circHipk3 on angiogenesis involved in cardiac regeneration[45]. The overexpression of circHipk3 promoted the proliferation, migration, and tube-forming capacity of coronary vessel endothelial cells. Enforced overexpression of circHipk3 alleviated cardiac dysfunction and fibrosis after MI in vivo. Mechanistically, circHipk3 enhanced cardiomyocyte proliferation by increasing N1ICD acetylation, and circHipk3 sponged miR-133a to promote CTGF expression in endothelial cells[45].
A study conducted by Li et al. revealed that statins upregulate circRNA-RBCK1, which is derived from the RBCK1 gene[46]. Moreover, statins increase AP-2α transcriptional activity, and AP-2α directly promotes RBCK1 expression in endothelial cells. Lovastatin protected diastolic function in mice, but this effect was abrogated by the absence of endothelial AP-2α or circRNA-RBCK1. Therefore, statins activate AP-2α/circRNA-RBCK1 signaling to block miR-133a activity and preserve diastolic function[46].
Recent data show that the abundance of hundreds of circRNAs is widely altered during TGF-β1-induced epithelial-mesenchymal transition (EMT), a cellular transition process similar to endothelial-mesenchymal transition (EndMT)[47-49]. However, the profiles of circRNAs during EndMT in cardiac remodeling and the functional outcomes of the related circRNAs influencing endothelial cell properties, cardiac remodeling and HF are still unknown.
Cardiac fibrosis
Cardiac fibrosis is a critical pathological feature of cardiac remodeling and HF and is driven by the activation of cardiac fibroblasts (CFs). The activation of CFs leads to the excessive deposition of extracellular matrix (ECM), resulting in stiffening of the chamber walls, impaired heart elasticity, impaired diastolic function and contractility, arrhythmias and local microfibrillations. This process is tightly regulated and characterized by increased synthesis of matrix metalloproteinases and stimulation of profibrotic mediators for the differentiation of CFs into myofibroblasts[50]. Different circRNAs have been reported to regulate cardiac fibrosis[51-67].
A study conducted by Wu et al. revealed the antifibrotic effect of circYAP in cardiac fibrosis[51]. CircYap was decreased in the myocardium of patients with myocardial hypertrophy and in mice subjected to TAC surgery. CircRNA ectopic expression in the mouse heart reduced the levels of fibrotic markers. The antifibrotic effects of circYap are mediated by the regulation of TMP4 and ACTG protein interactions, with subsequent inhibition of actin polymerization and cardiac fibrosis[51]. Recently, Du et al. reported that circNAB1 encoded the protein NAB1-356, which interacted with EGR1 to inhibit fibrosis and inflammation in the atrium. Moreover, NAB1-356 also suppressed the activities of Runx1 and Gadd45b, exerting inhibitory effects on cytokine expression and fibrosis[52].
Recently, Zhang et al. reported that circCAMTA1 expression is upregulated in the atrial appendage of patients with atrial fibrillation (AF)[53]. In fibroblasts isolated from the atrial appendage of patients with AF, Ang II treatment stimulated cell proliferation and the expression of fibrotic markers, which were mediated by increased levels of circCAMTA1. In vitro experiments demonstrated that the circCAMTA1-induced expression of fibrotic markers and proliferation were mediated by a reduction in the miR-214-3p expression and the activation of the TGFβR1 pathway. In a mouse model of Ang II-induced fibrosis, atrial interstitial fibrosis and both AF inducibility and durability were reduced, and fractional shortening was increased in the absence of circCAMTA1. These data indicate that the circCAMTA1/miR-214-3p/TGFβR1 axis regulates fibrosis-induced AF[53].
CircNlgn was increased in the myocardium of patients with selected congenital heart defects. CircNlgn encoded Nlgn173 with a 9-amino-acid nuclear localization motif, which combined with LaminB1 to facilitate the nuclear enrichment of Nlgn173. Nlgn173 promoted ING4 and C8orf44-SGK3 expression, contributing to aberrant collagen deposition and the activation of CFs. However, knockdown of circNlgn or ING4 and SGK3 expression abrogated cardiac fibrosis in mice with left ventricular pressure overload[54].
Mei et al. reported that the level of circSMAD3 was significantly reduced in murine HF models[55]. Overexpression of circSMAD3 inhibited cardiomyocyte hypertrophy and cardiac fibroblast activation. Mechanistically, circSMAD3 stabilized YBX1 and enhanced its interaction with SMAD3 in the nucleus to disrupt TGFβ/SMAD3 signaling[55].
Jung et al. reported that CircHIPK2 interacts with G3BP1 to promote stress granule formation in macrophages, initiating a cascade of downstream inflammatory events[56]. In contrast, circHIPK2 knockdown suppressed inflammatory signaling and inhibited proinflammatory cytokine secretion and cardiac fibrosis progression[56].
Mitochondrial dysfunction
Energy deficit leads to contractile dysfunction and alterations in cardiac energy metabolism, which is closely associated with the severity of pathological cardiac remodeling of the failing heart. An increase in ATP production from glycolysis and the alteration of fuels for mitochondrial ATP production are important ways to compensate for the cardiac energy deficit[68,69]. Metabolic alterations in the heart can cause changes in mitochondrial dynamics and Ca2+ homeostasis, leading to the overproduction of ROS and mitochondrial dysfunction. High levels of ROS can result in the accumulation of damaged DNA, proteins and lipids, thereby promoting inflammation, fibrosis and activation of the apoptotic pathway, ultimately leading to cell death and impairment of heart function[70]. Increasing evidence suggested various kinds of circRNAs are involved in mitochondrial dysfunction[71-75].
Zheng et al. identified circSamd4 as a mitochondrial-localized circRNA that regulates mitochondrial oxidative stress in CMs[71]. CircSamd4 overexpression reduced mitochondrial oxidative stress and oxidative DNA damage and induced CM proliferation after MI surgery. Mechanistically, circSamd4 increased the mitochondrial translocation of the Vcp protein, and Vcp decreased the Vdac1 level and prevented the opening of the mitochondrial permeability transition pore, revealing that circSamd4 is a potential target for HF therapy[71].
Circ-INSR, a circRNA derived from the insulin receptor gene, was significantly decreased in mouse and human failing heart tissue. It was also downregulated in patients who received anti-cancer treatment with doxorubicin. Circ-INSR knockdown resulted in impaired metabolic activity and in doxorubicin-treated CMs. Circ-INSR overexpression in the HL-1 murine cardiomyocyte cell line and primary rat CMs inhibited doxorubicin-induced apoptosis. The overexpression of circ-INSR improved doxorubicin-mediated myocardial dysfunction and atrophy, suggesting the potential of circ-INSR as a therapeutic candidate for doxorubicin-induced cardiotoxicity[31].
Recently, Li et al. reported that circSLC8a1 (hsa_circ_0005232) translated protein of SLC8a1-604[72]. HF phenotypes were exhibited in SLC8a1-604 transgenic mice but not in litter-matched negative controls. Mechanistically, SLC8a1-604 is translocated into mitochondria, where SLC8a1-604 combines with POLRMT, resulting in decreases in mitochondrial gene expression and ATP synthesis[72].
PYRCR was decreased in H/R-exposed cardiomyocytes and in the myocardium of I/R-injured mouse hearts. The overexpression of PYRCR inhibited cardiomyocyte pyroptosis, attenuated I/R-induced MI and improved cardiac function in mice. GTP-binding protein 2 (DRG2) was identified as the direct target of PYRCR. In addition, myocardial-specific DRG2 knockout enhanced the cardiac function of mice after I/R injury. DRG2 binds to dynamin-related protein 1 (Drp1) to increase its stability. In particular, PYRCR competitively abolished the interaction between DRG2 and Drp1, resulting in reduced mitochondrial fission and myocardial injury[73].
CIRCULATINg circRNAs AS BIOMARKERS OF CARDIAC REMODELING AND HF
Increasing evidence has shown that the aberrant expression of ncRNAs is associated with the pathophysiology of different conditions[76]. Therefore, ncRNA-based biomarkers, such as miRNAs and lncRNAs, have been proposed as innovative tools for the diagnosis and prognostication of cardiovascular diseases, including HF[77].
CircRNAs have specific properties that are useful in the field of clinical biomarkers; they are protected from degradation by exoribonucleases because of their covalently closed circular structure. Moreover, the diagnosis of cardiovascular diseases could be improved by combining the levels of circulating circRNAs with clinical information in an artificial intelligence prediction model[78].
Several circRNAs possess excellent biomarker potential in cardiac remodeling [Table 3]. The levels of circ_0112085, circ_0053919, circ_0062960 and circ_0014010 were markedly increased in the plasma of HF patients compared with those in the plasma of healthy controls. In addition, the level of circ_0062960 was positively associated with the serum level of B-type natriuretic peptide[79]. Particularly, Cdr1as was upregulated in the circulation of patients with chronic HF[80], and circ_0118464 was markedly increased in the epicardial adipose tissue of patients with HFpEF[81].
CircRNAs as biomarkers in cardiac remodeling and HF
| circRNA | Sample matrix | Expression change | Disease | Association | Reference |
| hsa_circ_0112085; hsa_circ_0062960; hsa_circ_0053919 | Plasma | Up | HF vs. healthy | hsa_circ_0062960 is positively associated with serum levels of BNP | [79] |
| Cdr1as | Plasma | Up | Chronic HF | miR-135a,b/HMOX1 | [80] |
| has_circ_0118464 | Epicardial adipose tissue | Up | HFpEF vs. non-HF control | With the highest fold-change | [81] |
| hsa_circ_0003258; hsa_circ_0051238; hsa_circ_0051239 | Plasma | Up | LMNA-related dilated cardiomyopathy vs. healthy | [82] | |
| hsa_circ_0089762 | Plasma | Up | Ischaemic DCM | Biomarkers for ischemic DCM | |
| hsa_circ_0097435 | Peripheral blood samples and EVs | UP | HF | As potential blood marker for HF | [83] |
| ACR | Plasma | Down | Chronic HF | ACR decrease positive with survival of HF | [84] |
| hsa_circ_0040414 | Plasma | Up | Chronic HF | miR-186-5p/Akt/PTEN | [85] |
| 9 hub dysregulated circRNAs | Plasma | Up | Chronic HF | 49 uniquely expressed circRNAs | [86] |
Four upregulated circRNAs were identified in the plasma of patients with dilated cardiomyopathy: circ_0003258, circ_0051239 and circ_0051238 in lamin A/C-related dilated cardiomyopathy and circ_0089762 in ischemic dilated cardiomyopathy[82]. Han et al. reported that circ_0097435 expression was significantly elevated in 40 HF patients. Circ_0097435 expression is increased in doxorubicin-treated cardiomyocytes, promoting cardiomyocyte apoptosis, indicating that circ_0097435 is a potential plasma marker of myocardial injury[83].
Circulating circRNAs are usually contained in extracellular vesicles (EVs), such as exosomes, microvesicles and apoptotic bodies, which can protect circRNA cargos from degradation[84-87]. RNA-seq of cardiac EVs from mouse hearts after I/R injury revealed that approximately 185 circRNAs were differentially expressed between these mice and Sham control mice[88]. The plasma exosomal RNA of 15 chronic coronary syndrome (CCS) patients and 15 noncardiac chest pain patients (NCCP, control group) was subjected to an RNA-seq assay to screen for dysregulated circRNAs. A total of 276 exosomal circRNAs were dysregulated in CCS patients. Among the 103 upregulated circRNAs, hsa_circ_0075269 and hsa_circ_0000284 are suggested to be potential biomarkers for CCS[89].
Moreover, the levels of circ_0097435 were higher not only in the plasma of patients with HF than in that of healthy subjects but also in exosomes isolated from these patients[82]. RNA sequencing revealed 893 dysregulated exosomal circRNAs in patients with AMI. Exosomal circ_0001558, circ_0001535, and circ_0000972 were significantly upregulated in AMI patients. In addition, the expression of circ_0001558 increased 4.45-fold in a second cohort of AMI patients, indicating that it is a promising diagnostic indicator for AMI[90].
These findings motivate further characterization of the potential of circulating circRNAs encapsulated in EVs as indicators of pathological cardiac remodeling and HF. Accurate and sensitive quantification of exosomal circRNAs is critical for their application as clinical biomarkers. In addition to reverse transcription quantitative PCR (RT-qPCR), digital PCR (dPCR) technology has recently emerged as a new-generation platform for exosomal circRNA quantification. However, the use of exosomal circRNAs as diagnostic markers for cardiac remodeling and HF has several limitations, such as the isolation and characterization of exosomes and methods for the quantification of circRNAs. Currently, commercial kits are used to extract exosomes from blood, but the purity of these exosomes does not meet the minimal information for diagnostic evaluation[91]. The number of clinical samples should be expanded to validate the diagnostic efficacy of the potential exosomal circRNA biomarkers for cardiac remodeling and HF. Moreover, further investigations are necessary to evaluate biomarker performance characteristics, such as sensitivity, specificity and accuracy, and compare biomarker values with existing indicators. In the future, dPCR combined with machine learning will be an important strategy for the analysis of exosomal circRNAs for cardiac remodeling and HF diagnosis.
THERAPEUTIC POTENTIAL AND PERSPECTIVE OF circRNAs IN HF
To date, circRNA-based approaches have been investigated in preclinical studies. Small interfering RNA (siRNA) targeting the junction site sequence of circRNA is a commonly used strategy to knock down circRNA expression. However, enhancing the stability of siRNA, optimizing the targeted delivery of siRNA to cardiac cells and minimizing off-target effects of the designed siRNA are fundamental. Compared with an RNAi-based approach, the CRISPR/Cas13 system could suppress circRNAs more efficiently and with lower off-target effects, but its effectiveness in vivo warrants further investigation[92]. Moreover, the delivery system of CRISPR/Cas13 and the side effects of Cas13 expression in vivo are elusive and required further demonstration.
Lentiviruses containing instructions for generating circRNAs are typically used for circRNA overexpression in vivo[93]. Synthetic circRNAs can also be used to administer circRNA. The engineering of artificial circRNAs affecting interactions with detrimental miRNAs and RBPs is a potential future therapeutic approach for HF. CircRNA sponges (circmiRs) with bulged binding sites for prohypertrophic miRNAs, such as miR-132-3p and -212-3p, were expressed via AAV9 vectors in the myocardium of TAC mice, and myocardial hypertrophy was attenuated, and cardiac function was preserved[94]. Similarly, artificial circSP199a includes complements of the seed sequence of miR-199a-5p, and -3p was designed for functional studies. Exogenous administration of circSP199a, specifically sponges miR-199a-5p and -3p, results in the amelioration of myocardial hypertrophy and fibrosis in TAC mice[95]. Moreover, artificial circRNAs can be designed to carry IRES and ORF elements for protein translation, which could be a viable option for therapeutic protein expression[96]. However, artificial circRNAs are not widely used to mimic endogenous circRNAs. Currently, the technology of circRNA chemical synthesis and the enhancement of their protein compilation capability require further improvement and development.
Despite the therapeutic potential of circRNAs, their stability, half-life, immunogenicity and delivery routes to ensure cardiac uptake and additional pharmacological issues remain to be determined. Synthetic circRNAs are homogeneous and do not elicit an immune response in vivo. Moreover, a suitable delivery system for exogenous circRNA can transport circRNAs to cells and enable them to evade surveillance by the autoimmune system.
Smaller liposomes are known to have greater abilities to escape phagocyte uptake, penetrate targeted cells and ameliorate the risk of systemic toxicity[97]. Stabilized lipid-based nanoparticles (LNPs) are a new kind of liposome that include solid lipid nanoparticles, nanostructured lipid carriers, and cationic lipid amphiphiles and possess improved internal physical stability[98,99]. LNPs, as promising vehicles, have emerged across the biomedical industry to deliver therapeutic circRNA drugs. However, the distribution and expression kinetics of circRNAs depend on the optimization of lipid components, LNP adjuvants and routes of administration[100,101].
CONCLUSION
CircRNAs are recognized as specific types of lncRNAs that participate in the pathogenesis of cardiac remodeling and HF. CircRNAs play critical roles in regulating gene expression at the transcriptional, posttranscriptional, and translational levels. The regulatory function of circRNAs in cardiac remodeling and HF mechanisms, including cardiomyocyte death, cardiac hypertrophy, fibrosis, EC dysfunction and functional disorders of mitochondria, has received increasing attention and in-depth development in recent years.
CircRNAs possess unique properties and are promising candidates for translational medicine research in cardiac remodeling and HF. CircRNAs as potential HF biomarkers require the persistent optimization of standard operating techniques and a reduction in costs for detection in clinics. From the perspective of therapeutic drugs, effective strategies focused on the precise molecular design and heart-targeted delivery of circRNAs need to be developed, as well as eliminating off-target effects and undesired immune responses. Further investigations involving mechanistic research, technological innovation, and translational studies of circRNA drugs will improve the therapeutic potential of these drugs in combating cardiac remodeling and HF.
DECLARATIONS
Authors’ contributions
Collected the materials and drafted the manuscript: Li R, Su X
Collected the materials and prepared the figures: Jiang J, Su J
Prepared the tables: Zhu J, Xu J
Designed the study and revised the manuscript: Shan Z
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported by the National Natural Science Foundation of China (No. 92581127, No.82570331, No. 82470253, No. 82300277) and Guangdong Basic and Applied Basic Research Foundation (No. 2026A1515010884, No. 2025A15150112490, No. 2025A1515010873).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
2. Timmis A, Townsend N, Gale CP, et al. European society of cardiology: cardiovascular disease statistics 2019. Eur Heart J. 2020;41:12-85.
3. Perrino C. Intermittent pressure overload triggers hypertrophy-independent cardiac dysfunction and vascular rarefaction. J Clin Investig. 2006;116:1547-60.
4. Frantz S, Hundertmark MJ, Schulz-Menger J, Bengel FM, Bauersachs J. Left ventricular remodelling post-myocardial infarction: pathophysiology, imaging, and novel therapies. Eur Heart J. 2022;43:2549-61.
5. Schirone L, Forte M, D’ambrosio L, et al. An overview of the molecular mechanisms associated with myocardial ischemic injury: state of the art and translational perspectives. Cells. 2022;11:1165.
6. Micheel J, Safrastyan A, Wollny D. Advances in non-coding RNA sequencing. Noncoding RNA. 2021;7:70.
7. Gao J, Xu W, Wang J, Wang K, Li P. The role and molecular mechanism of non-coding RNAs in pathological cardiac remodeling. Int J Mol Sci. 2017;18:608.
8. Jha S, Thasma Loganathbabu VK, Kumaran K, Krishnasamy G, Aruljothi KN. Long non-coding RNAs (lncRNAs) in heart failure: a comprehensive review. Noncoding RNA. 2023;10:3.
9. Shen S, Jiang H, Bei Y, Xiao J, Li X. Long non-coding RNAs in cardiac remodeling. Cell Physiol Biochem. 2017;41:1830-7.
10. Salzman J, Gawad C, Wang PL, Lacayo N, Brown PO. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE. 2012;7:e30733.
11. Zhang C, Huo ST, Wu Z, et al. Rapid development of targeting circRNAs in cardiovascular diseases. Mol Ther Nucleic Acids. 2020;21:568-76.
12. Ding C, Zhou Y. Insights into circular RNAs: biogenesis, function and their regulatory roles in cardiovascular disease. J Cell Mol Med. 2023;27:1299-314.
13. Joaquim VHA, Pereira NP, Fernandes T, Oliveira EM. Circular RNAs as a diagnostic and therapeutic target in cardiovascular diseases. Int J Mol Sci. 2023;24:2125.
14. Wang X, Shan G. Features and biomedical relevance of circular RNA biogenesis. Trends Genet. 2026;42:282-94.
15. Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495:384-8.
16. Zhou W, Cai Z, Liu J, Wang D, Ju H, Xu R. Circular RNA: metabolism, functions and interactions with proteins. Mol Cancer. 2020;19:172.
17. Hwang HJ, Kim YK. Molecular mechanisms of circular RNA translation. Exp Mol Med. 2024;56:1272-80.
18. Capirossi G, Brasini S, Tremoli E, Binatti A, Roncarati R. Circular RNAs in cardiovascular physiopathology: from molecular mechanisms to therapeutic opportunities. Int J Mol Sci. 2025;26:9725.
20. Geng H, Li R, Su Y, et al. The circular RNA Cdr1as promotes myocardial infarction by mediating the regulation of miR-7a on its target genes expression. PLoS ONE. 2016;11:e0151753.
21. Wang L, Yu P, Wang J, et al. Downregulation of circ-ZNF609 promotes heart repair by modulating RNA N6-methyladenosine-modified Yap expression. Research. 2022;2022:9825916.
22. Yu P, Wang J, Xu G, et al. RNA m6A-regulated circ-ZNF609 suppression ameliorates doxorubicin-induced cardiotoxicity by upregulating FTO. JACC Basic Transl Sci. 2023;8:677-98.
23. Zhu Y, Zhao P, Sun L, et al. Overexpression of circRNA SNRK targets miR-103-3p to reduce apoptosis and promote cardiac repair through GSK3β/β-catenin pathway in rats with myocardial infarction. Cell Death Discov. 2021;7:84.
24. Yuan Q, Sun Y, Yang F, et al. CircRNA DICAR as a novel endogenous regulator for diabetic cardiomyopathy and diabetic pyroptosis of cardiomyocytes. Sig Transduct Target Ther. 2023;8:99.
25. Wang D, Tian L, Wang Y, Gao X, Tang H, Ge J. Circ_0001206 regulates miR-665/CRKL axis to alleviate hypoxia/reoxygenation-induced cardiomyocyte injury in myocardial infarction. ESC Heart Fail. 2022;9:998-1007.
26. Li Y, Ren S, Xia J, Wei Y, Xi Y. EIF4A3-induced circ-BNIP3 aggravated hypoxia-induced injury of H9c2 cells by targeting miR-27a-3p/BNIP3. Mol Ther Nucleic Acids. 2020;19:533-45.
27. Li D, You J, Mao C, et al. Circular RNA Fbxl5 regulates cardiomyocyte apoptosis during ischemia reperfusion injury via sponging microRNA-146a. J Inflamm Res. 2022;15:2539-50.
28. Wang L, Feng J, Feng X, et al. Exercise-induced circular RNA circUtrn is required for cardiac physiological hypertrophy and prevents myocardial ischaemia-reperfusion injury. Cardiovasc Res. 2023;119:2638-52.
29. Gao X, Liu C, Zhang Y, et al. The circRNA CNEACR regulates necroptosis of cardiomyocytes through Foxa2 suppression. Cell Death Differ. 2021;29:527-39.
30. Ju J, Li X, Zhao X, et al. Circular RNA FEACR inhibits ferroptosis and alleviates myocardial ischemia/reperfusion injury by interacting with NAMPT. J Biomed Sci. 2023;30:45.
31. Lu D, Chatterjee S, Xiao K, et al. A circular RNA derived from the insulin receptor locus protects against doxorubicin-induced cardiotoxicity. Eur Heart J. 2022;43:4496-511.
32. Shen M, Wu Y, Li L, Zhang L, Liu G, Wang R. CircMAP3K5 promotes cardiomyocyte apoptosis in diabetic cardiomyopathy by regulating miR‐22‐3p/DAPK2 Axis. J Diabetes. 2023;16:e13471.
33. Pitoulis FG, Terracciano CM. Heart plasticity in response to pressure- and volume-overload: a review of findings in compensated and decompensated phenotypes. Front Physiol. 2020;11:92.
34. Shimizu I, Minamino T. Physiological and pathological cardiac hypertrophy. J Mol Cell Cardiol. 2016;97:245-62.
35. Li M, Ding W, Fang X, et al. Novel truncated peptide derived from circCDYL exacerbates cardiac hypertrophy. Circ Res. 2025;136:e94-112.
36. Pan W, Hunkler HJ, Chatterjee S, et al. A circular RNA derived from the ryanodine receptor 2 locus controls cardiac hypertrophy and calcium handling. Cell Mol Life Sci. 2025;82:359.
37. Yang M, Wang W, Wang L, Li Y. Circ_0001052 promotes cardiac hypertrophy via elevating Hipk3. Aging. 2023;15:204521.
38. Wang K, Long B, Liu F, et al. A circular RNA protects the heart from pathological hypertrophy and heart failure by targeting miR-223. Eur Heart J. 2016;37:2602-11.
39. Xu Q, Liu J, Zhu R, et al. NSD2 promotes pressure overload-induced cardiac hypertrophy via activating circCmiss1/TfR1/ferroptosis signaling. Life Sci. 2023;328:121873.
40. Pan J, Xu Z, Guo G, et al. Circ_nuclear factor I X (circNfix) attenuates pressure overload-induced cardiac hypertrophy via regulating miR-145-5p/ATF3 axis. Bioengineered. 2021;12:5373-85.
41. Li H, Xu J, Fang X, et al. Circular RNA circRNA_000203 aggravates cardiac hypertrophy via suppressing miR-26b-5p and miR-140-3p binding to Gata4. Cardiovasc Res. 2020;116:1323-34.
42. Lin X, Zhang L, Zhang W, Lei X, Lu Q, Ma A. Circular RNA circ_0001006 aggravates cardiac hypertrophy via miR-214-3p/PAK6 axis. Aging. 2022;14:2210-20.
43. Miličić D, Jakuš N, Fabijanović D. Microcirculation and heart failure. Curr Pharm Des. 2018;24:2954-9.
44. Vancheri F, Longo G, Vancheri S, Henein M. Coronary microvascular dysfunction. J Clin Med. 2020;9:2880.
45. Si X, Zheng H, Wei G, et al. circRNA Hipk3 induces cardiac regeneration after myocardial infarction in mice by binding to notch1 and miR-133a. Mol Ther Nucleic Acids. 2020;21:636-55.
46. Li B, Bai W, Guo T, et al. Statins improve cardiac endothelial function to prevent heart failure with preserved ejection fraction through upregulating circRNA-RBCK1. Nat Commun. 2024;15:2953.
47. Conn SJ, Pillman KA, Toubia J, et al. The RNA binding protein quaking regulates formation of circRNAs. Cell. 2015;160:1125-34.
48. Liao J, Wu J, Wang Y, et al. Deep sequencing reveals a global reprogramming of lncRNA transcriptome during EMT. Biochim Biophys Acta Mol Cell Res. 2017;1864:1703-13.
49. Madè A, Bibi A, Garcia-Manteiga JM, et al. circRNA-miRNA-mRNA deregulated network in ischemic heart failure patients. Cells. 2023;12:2578.
50. Yamada Y, Sadahiro T, Nakano K, et al. Cardiac reprogramming and Gata4 overexpression reduce fibrosis and improve diastolic dysfunction in heart failure with preserved ejection fraction. Circulation. 2025;151:379-95.
51. Wu N, Xu J, Du WW, et al. YAP circular RNA, circYap, attenuates cardiac fibrosis via binding with tropomyosin-4 and gamma-actin decreasing actin polymerization. Mol Ther. 2021;29:1138-50.
52. Du WW, Rafiq M, Yuan H, et al. A novel protein NAB1‐356 encoded by circRNA circNAB1 mitigates atrial fibrillation by reducing inflammation and fibrosis. Adv Sci. 2025;12:2411959.
53. Zhang L, Lou Q, Zhang W, et al. CircCAMTA1 facilitates atrial fibrosis by regulating the miR-214-3p/TGFBR1 axis in atrial fibrillation. J Mol Histol. 2022;54:55-65.
54. Du WW, Xu J, Yang W, et al. A neuroligin isoform translated by circNlgn contributes to cardiac remodeling. Circ Res. 2021;129:568-82.
55. Mei S, Ma X, Zhou L, et al. CircSMAD3 represses SMAD3 phosphorylation and ameliorates cardiac remodeling by recruiting YBX1. iScience. 2024;27:111305.
56. Jung M, Schmidt A, Sansonetti M, et al. Macrophage-specific circular RNA circHIPK2, inflammation, and fibrosis after myocardial infarction. Eur Heart J. 2026;47:2831-47.
57. Jeong A, Lim Y, Kook T, et al. Circular RNA circSMAD4 regulates cardiac fibrosis by targeting miR-671-5p and FGFR2 in cardiac fibroblasts. Mol Ther Nucleic Acids. 2023;34:102071.
58. Wang Y, Wang H, Zhang L, Zhang J, Liu N, Zhao P. A novel identified circular RNA, circSnap47, promotes heart failure progression via regulation of miR-223-3p/MAPK axis. Mol Cell Biochem. 2022;478:459-69.
59. Schmidt A, Schmidt K, Groß S, et al. Circular RNA circIGF1R controls cardiac fibroblast proliferation through regulation of carbohydrate metabolism. Sci Rep. 2025;15:20331.
60. Bibi A, Madè A, Greco S, et al. Circular PVT1 promotes cardiac fibroblast activation interacting with miR-30a-5p and miR-125b-5p. Cell Death Dis. 2025;16:325.
61. Yan J, Ai C, Chen Q, et al. CircMap4k2 reactivated by aneurysm plication alleviates residual cardiac remodeling after SVR by enhancing cardiomyocyte proliferation in post-MI mice. J Adv Res. 2024;65:227-38.
62. Ma GB, Chen WX, Zhan FJ, et al. Circ_0002295 facilitated myocardial fibrosis progression through the miR‐1287/CXCR2 axis. Clin Exp Pharmacol Physiol. 2023;50:944-53.
63. Wang Y, Li C, Zhao R, et al. CircUbe3a from M2 macrophage-derived small extracellular vesicles mediates myocardial fibrosis after acute myocardial infarction. Theranostics. 2021;11:6315-33.
64. Jiang Y, Zhang B, Zhang B, et al. A novel feedback loop: CELF1/circ-CELF1/BRPF3/KAT7 in cardiac fibrosis. Acta Pharm Sin B. 2025;15:5192-211.
65. Guo J, Chen L, Huang Z, et al. Suppression of the inhibitory effect of circ_0036176-translated Myo9a-208 on cardiac fibroblast proliferation by miR-218-5p. J Cardiovasc Transl Res. 2022;15:548-59.
66. Pang P, Si W, Wu H, et al. The circular RNA circHelz enhances cardiac fibrosis by facilitating the nuclear translocation of YAP1. Transl Res. 2023;257:30-42.
67. Sun LY, Zhao JC, Ge XM, Zhang H, Wang CM, Bie ZD. Circ_LAS1L regulates cardiac fibroblast activation, growth, and migration through miR‐125b/SFRP5 pathway. Cell Biochem Funct. 2020;38:443-50.
68. Lopaschuk GD, Karwi QG, Tian R, Wende AR, Abel ED. Cardiac energy metabolism in heart failure. Circ Res. 2021;128:1487-513.
69. Sun Q, Karwi QG, Wong N, Lopaschuk GD. Advances in myocardial energy metabolism: metabolic remodelling in heart failure and beyond. Cardiovasc Res. 2024;120:1996-2016.
70. Gibb AA, Hill BG. Metabolic coordination of physiological and pathological cardiac remodeling. Circ Res. 2018;123:107-28.
71. Zheng H, Huang S, Wei G, et al. CircRNA Samd4 induces cardiac repair after myocardial infarction by blocking mitochondria-derived ROS output. Mol Ther. 2022;30:3477-98.
72. Li F, Du WW, Li X, et al. A new protein isoform encoded by human circular RNA circSLC8a1 contributes to cardiac remodelling. Cardiovasc Res. 2025;121:882-99.
73. Chen X, Xu H, Zhao X, et al. PYRCR alleviates myocardial ischemia/reperfusion injury in mice via inhibiting DRG2-mediated cardiomyocyte pyroptosis. Acta Pharmacol Sin. 2025;46:3204-16.
74. Ni X, Duan L, Bao Y, et al. Circ_005077 accelerates myocardial lipotoxicity induced by high-fat diet via CyPA/p47PHOX mediated ferroptosis. Cardiovasc Diabetol. 2024;23:129.
75. Wang Z, Liu X, Deng Y. Negative feedback of SNRK to circ-SNRK regulates cardiac function post-myocardial infarction. Cell Death Differ. 2021;29:709-21.
76. Juni RP, Phelp PG, Bink DI, et al. COVID19 impairs cardiac function via endothelial H19 and IL6 signaling. Circ Res. 2023;132:648-51.
77. Robinson EL, Baker AH, Brittan M, et al. Dissecting the transcriptome in cardiovascular disease. Cardiovasc Res. 2022;118:1004-19.
78. Zhou Q, Boeckel JN, Yao J, et al. Diagnosis of acute myocardial infarction using a combination of circulating circular RNA cZNF292 and clinical information based on machine learning. MedComm. 2023;4:e299.
79. Sun Y, Jiang X, Lv Y, et al. Circular RNA expression profiles in plasma from patients with heart failure related to platelet activity. Biomolecules. 2020;10:187.
80. Chen C, Shen H, Huang Q, Li Q. The circular RNA CDR1as regulates the proliferation and apoptosis of human cardiomyocytes through the miR-135a/HMOX1 and miR-135b/HMOX1 axes. Genet Test Mol Biomarkers. 2020;24:537-48.
81. He S, Zhu H, Zhang J, Yang X, Zhao L. Genome-wide screening for circRNAs in epicardial adipose tissue of heart failure patients with preserved ejection fraction. Am J Transl Res. 2023;15:4610-9.
82. Costa MC, Calderon-Dominguez M, Mangas A, et al. Circulating circRNA as biomarkers for dilated cardiomyopathy etiology. J Mol Med. 2021;99:1711-25.
83. Han J, Zhang L, Hu L, et al. Circular RNA-expression profiling reveals a potential role of Hsa_circ_0097435 in heart failure via sponging multiple MicroRNAs. Front Genet. 2020;11:212.
84. Yan H, Du D, Wang C, Tian M. Downregulation of autophagy-related circular RNA (ACR) is correlated with poor survival of patients with chronic heart failure. Bioengineered. 2022;13:13141-9.
85. Feng Y, Yan B, Cheng H, et al. Knockdown circ_0040414 inhibits inflammation, apoptosis and promotes the proliferation of cardiomyocytes via miR‐186‐5p/PTEN/AKT axis in chronic heart failure. Cell Biol Int. 2021;45:2304-15.
86. Xu S, Zhang G, Tan X, et al. Differential expression spectrum of circRNA in plasma exosomes in dilated cardiomyopathy with heart failure. J Cell Mol Med. 2024;28:e70258.
87. Collado A, Gan L, Tengbom J, Kontidou E, Pernow J, Zhou Z. Extracellular vesicles and their non‐coding RNA cargos: emerging players in cardiovascular disease. J Physiol. 2022;601:4989-5009.
88. Ge X, Meng Q, Zhuang R, et al. Circular RNA expression alterations in extracellular vesicles isolated from murine heart post ischemia/reperfusion injury. Int J Cardiol. 2019;296:136-40.
89. Liu X, Zheng M, Han R, et al. Circulating exosomal CircRNAs as diagnostic biomarkers for chronic coronary syndrome. Metabolites. 2023;13:1066.
90. Liu X, Zhang Y, Yuan W, et al. Exosomal CircRNAs in circulation serve as diagnostic biomarkers for acute myocardial infarction. Front Biosci. 2024;29:149.
91. Yang D, Zhang W, Zhang H, et al. Progress, opportunity, and perspective on exosome isolation - efforts for efficient exosome-based theranostics. Theranostics. 2020;10:3684-707.
92. Li S, Li X, Xue W, et al. Screening for functional circular RNAs using the CRISPR-Cas13 system. Nat Methods. 2020;18:51-9.
93. Holdt LM, Kohlmaier A, Teupser D. Circular RNAs as therapeutic agents and targets. Front Physiol. 2018;9:1262.
94. Lavenniah A, Luu TDA, Li YP, et al. Engineered circular RNA sponges act as miRNA inhibitors to attenuate pressure overload-induced cardiac hypertrophy. Mol Ther. 2020;28:1506-17.
95. Wu H, Zhou C, Gao Y, et al. circSP199a, a circularized RNA sponge targeting miR-199a-5p and -3p, mitigates mouse cardiac hypertrophy and fibrosis. Acta Pharmacol Sin. 2025;47:86-102.
96. Wesselhoeft RA, Kowalski PS, Anderson DG. Engineering circular RNA for potent and stable translation in eukaryotic cells. Nat Commun. 2018;9:2629.
97. Inglut CT, Sorrin AJ, Kuruppu T, et al. Immunological and toxicological considerations for the design of liposomes. Nanomaterials. 2020;10:190.
98. Tenchov R, Bird R, Curtze AE, Zhou Q. Lipid nanoparticles-from liposomes to mRNA vaccine delivery, a landscape of research diversity and advancement. ACS Nano. 2021;15:16982-7015.
99. Niu D, Wu Y, Lian J. Circular RNA vaccine in disease prevention and treatment. Sig Transduct Target Ther. 2023;8:341.
100. Qiu M, Li Y, Bloomer H, Xu Q. Developing biodegradable lipid nanoparticles for intracellular mRNA delivery and genome editing. Acc Chem Res. 2021;54:4001-11.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0











Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].