


The role of noncoding RNA in aortic dissection
 0
0 Abstract
Aortic dissection (AD) is a rapidly progressive cardiovascular disease associated with high mortality. The core pathogenic mechanisms of AD include apoptosis and phenotypic switching of vascular smooth muscle cells (VSMCs), remodeling and degradation of the extracellular matrix (ECM), and vascular inflammation. However, the detailed molecular mechanisms remain incompletely understood. Noncoding RNAs (ncRNAs) are RNA molecules that do not encode proteins. Recent studies have confirmed that ncRNAs with aberrant expression exert critical regulatory functions in the pathogenesis and progression of AD, and several ncRNAs have shown significant application potential in the early diagnosis, conservative treatment, and prognostic assessment of AD. This review summarizes the latest research progress on the regulatory mechanisms of various ncRNAs in AD, and critically evaluates their potential clinical utility as clinical biomarkers and therapeutic targets for AD.
Keywords
INTRODUCTION
Aortic dissection (AD) is a cardiovascular disease characterized by sudden onset, rapid progression, and extremely high mortality. According to the 2022 ACC/AHA Guideline for the Diagnosis and Management of Aortic Disease, AD is defined as a pathological condition in which blood penetrates into the aortic media through a tear in the aortic intima, leading to the separation of the elastic lamellae and smooth muscle cell layer of the media. This tear spreads along the longitudinal axis of the aorta, forming an abnormal cavity (false lumen) parallel to the original vascular lumen (true lumen)[1]. This process can lead to the destruction of the aortic wall structure and impaired blood supply to branch vessels, which can induce fatal complications such as myocardial infarction, stroke, visceral ischemia, and aortic rupture[2]. The pathological essence of AD lies in the destruction of the structural integrity of the aortic wall, involving the abnormal phenotypic switching of vascular smooth muscle cells (VSMCs), excessive degradation of the extracellular matrix (ECM), and the involvement of sustained inflammatory and immune responses. These changes ultimately result in a significant decrease in the mechanical strength and elasticity of the aortic wall, rendering it susceptible to tearing and dilatation under hemodynamic stress[3-5].
According to the 2023 Report on Cardiovascular Health and Diseases in China, AD accounts for 48% of aortic diseases among Chinese residents[6]. Based on the extent of involvement, AD is classified into Stanford type A (including type I and II) and type B[7]. Data show that the mortality rate of patients with untreated type A AD increases by 1%-2% per hour after onset, reaching 33% at 48 h, over 50% at 1 week, and over 80% at 1 month[8]. Even for type B AD, which has a relatively better prognosis, it still poses a serious threat to the patients’ lives if not treated in a timely manner[9]. The high mortality rate of AD not only brings devastating blows to patients’ families but also imposes a heavy burden on social medical resources[10].
Noncoding RNAs (ncRNAs) are a class of RNA molecules that do not encode proteins or only encode short peptides after transcription, constituting the vast majority of the genome transcriptome[11]. According to molecular size and structural characteristics, ncRNAs are mainly divided into the following categories:
MicroRNAs (miRNAs): ~20-25 nucleotides in length. They are key molecules for rapid and precise gene regulation, which act by forming incomplete complementary binding with the 3’ untranslated region (3’UTR) of target messenger RNAs (mRNAs), thereby inducing mRNA degradation or translational repression[12].
Long noncoding RNAs (lncRNAs): over 200 nucleotides in length, with diverse structures and complex functions. They exert regulatory effects at the epigenetic, transcriptional, and post-transcriptional levels through multiple mechanisms, including chromatin modification, transcriptional interference, acting as competing endogenous RNA (ceRNA) “sponges” to sequester miRNAs, and interacting with proteins[13].
Circular RNAs (circRNAs): a class of closed-loop ncRNAs with high stability and conservation. One of their main functions is to act as highly efficient miRNA “sponges”, which relieve the inhibitory effect of miRNAs on target genes, thereby participating in various pathophysiological processes[14].
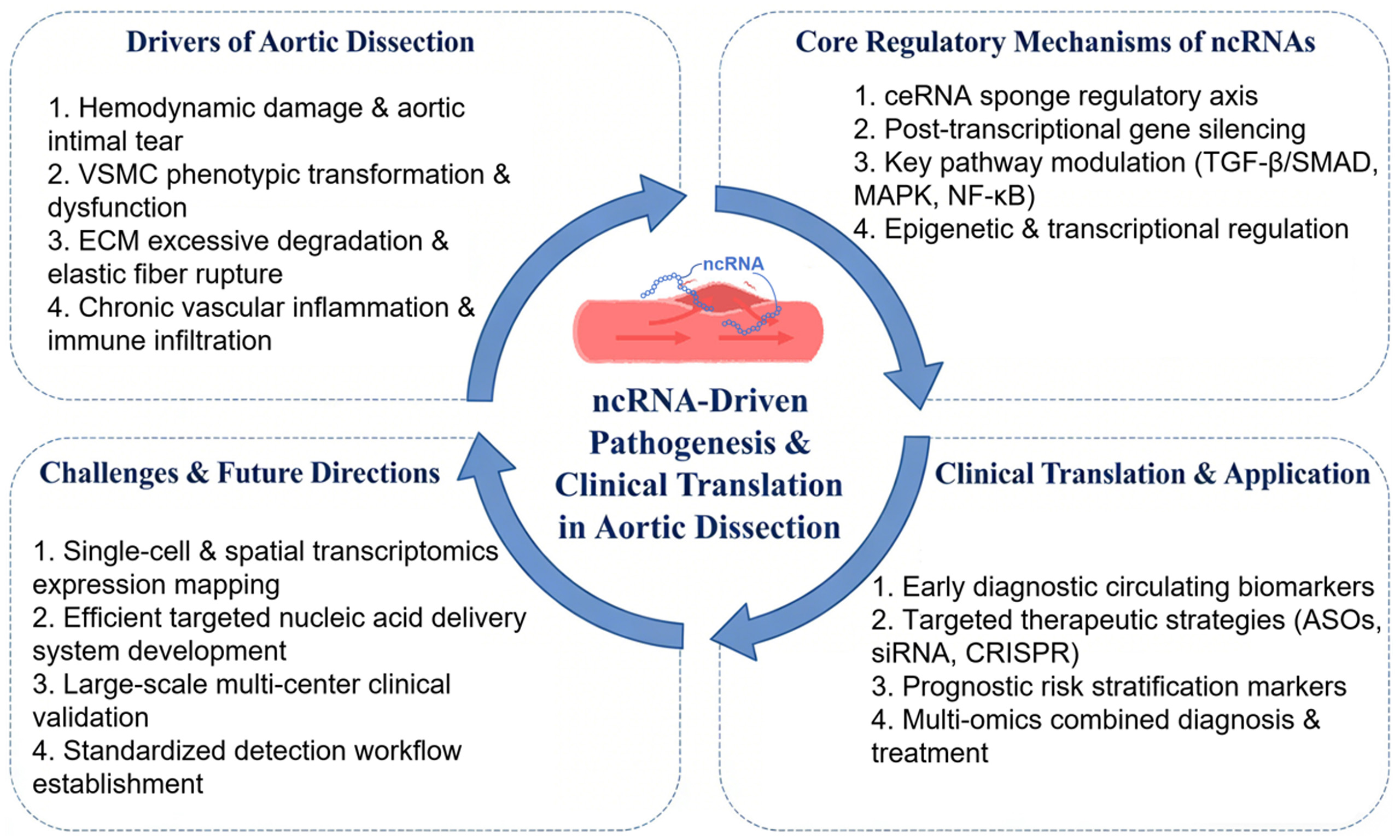
In recent years, a growing body of evidence has demonstrated that ncRNAs affect the pathogenesis and progression of AD by regulating the proliferation/phenotypic switching of VSMCs, inflammatory activation, and ECM remodeling through mechanisms such as post-transcriptional repression, sponge activity, and signaling pathway regulation[15]. Early research in this field focused predominantly on the regulatory roles of miRNAs in AD pathogenesis and progression. lncRNAs have more recently emerged as a major research hotspot, while circRNA research remains in its early exploratory phase. This review systematically delineates the functional mechanisms of distinct ncRNA subtypes in AD, outlines their translational progress as diagnostic biomarkers and therapeutic targets, discusses existing challenges in clinical translation, and highlights future research directions in this field.
PATHOPHYSIOLOGICAL MECHANISMS OF NCRNAS IN AD
Dysregulation of vascular smooth muscle cell function and its regulation by ncRNAs
As the principal cellular component of the aortic media, VSMCs and their functional dysregulation constitute the central driver of AD pathogenesis. During disease progression, ncRNAs exert their effects mainly by regulating key biological processes of VSMCs, including phenotypic switching, proliferation, apoptosis, and migration[16]. Among these, abnormal phenotypic switching is the core regulatory link, which is mainly achieved through key signaling pathways such as Transforming Growth Factor β (TGF-β), Mitogen-Activated Protein Kinase (MAPK), and Nuclear Factor-kappa B (NF-κB)[17,18].
ncRNAs (miRNAs, lncRNAs, and circRNAs) regulate signal transduction by targeting key molecules within these pathways - including receptors, kinases, inhibitory proteins, transcription factors - thus determining whether VSMCs maintain a normal contractile phenotype or switch to a pathogenic synthetic phenotype. Specifically, the TGF-β/SMAD (Sma/Mothers against decapentaplegic) axis is the core pathway for phenotypic regulation: ncRNAs regulate pathway activation and phenotypic switching by targeting TGF-β ligands, receptors, or SMAD family molecules (e.g. miR-145 targets and inhibits SMAD3[19]). The MAPK pathway (ERK1/2, JNK, p38) mediates the synthetic phenotype, as well as proliferation and migration: ncRNAs exert regulatory effects by targeting key kinases of the pathway (e.g. miR-22 targets and inhibits p38[20]). The NF-κB pathway links inflammation and phenotypic switching: ncRNAs target its key molecules to activate the pathway and upregulate pro-inflammatory factors, synergistically promoting the synthetic phenotype. Some ncRNAs can coordinately regulate multiple pathways, ultimately leading to the degradation of elastic fibers in the aortic media, impairment of vascular wall structure and function, and driving the progression of AD. The roles of ncRNAs in VSMCs are summarized in Table 1.
ncRNAs influencing AD development by regulating VSMC functions
| ncRNA type | Expression status | Target/pathway | Primary regulatory process | Role in AD | Potential application directions | References |
| miR-145 | Downregulation | SMAD3 | Promotes VSMC proliferation, migration, and apoptosis | Inhibits AD formation | Therapeutic target | [19] |
| miR-22 | Downregulation | p38, MAPK | Inhibits VSMC apoptosis | Inhibits AD formation | Therapeutic target | [20] |
| miR-21 | Upregulation | PTEN, SV2C | Promotes VSMC proliferation | Promotes AD development | Therapeutic target | [21] |
| miR-107-5p | Upregulation | ITM2C | Promotes VSMC proliferation | Promotes AD development | Therapeutic target | [22] |
| miR-124 | Downregulation | Sp1 | Promotes VSMC phenotypic switching | Inhibits AD formation | Therapeutic target | [23] |
| miR-134-5p | Downregulation | - | Inhibits VSMC phenotypic switching and migration | Inhibits AD formation | Therapeutic target | [24] |
| miR-200c-3p | Upregulation | CASZ1, Rho A/Egfl7 | Promotes VSMC phenotypic switching | Promotes AD development | Therapeutic target | [25] |
| lncRNA H19 | Upregulation | miR-193b-3p | Promotes VSMC phenotypic switching | Promotes AD development | Diagnostic/therapeutic target | [26] |
| lncRNA SENCR | Downregulation | miR-206 | Maintains VSMC contractile phenotype | Inhibits AD formation | Therapeutic target | [27] |
| lncRNA-HIF1A-AS2 | Upregulation | miR-33b | Promotes VSMC phenotypic switching | Promotes AD development | Therapeutic target | [28] |
| lncRNA-PVT1 | Upregulation | miR-27b-3p | Promotes VSMC phenotypic switching | Promotes AD development | Therapeutic target | [29] |
| lncRNA-CDKN2B-AS1 | Upregulation | miR-320d, STAT3 | Promotes VSMC apoptosis | Promotes AD development | Therapeutic target | [30] |
| lncRNA-PTENP1 | Upregulation | miR-21, Akt | Promotes VSMC apoptosis | Promotes AD development | Diagnostic/therapeutic target | [31] |
| lncRNA-XIST | Upregulation | miR-17 | Promotes VSMC apoptosis | Promotes AD development | Therapeutic target | [32] |
| lncRNA-LINC01278 | Downregulation | miR-500b-5p | Maintains VSMC contractile phenotype | Inhibits AD formation | Therapeutic target | [33] |
| lncRNA-RP11-465L10.10 | Upregulation | NF-κB | Promotes VSMC phenotypic switching Promotes ECM degradation | Promotes AD development | Diagnostic/therapeutic target | [34] |
| circ_0000006circ_0000160 | Upregulation | hsa-let-7e-5p, UBQLN4 | Promotes VSMC phenotypic switching | Promotes AD development | Diagnostic/therapeutic target | [35] |
| circ_0022920 | Downregulation | miR-650 | Inhibits VSMC proliferation and migration | Inhibits AD formation | Therapeutic target | [36] |
| circ-NRIP1 | Upregulation | IGF2BP1, CXCL5 | Promotes VSMC proliferation and migration | Promotes AD development | Diagnostic/therapeutic target | [37] |
| circ-TGFBR2 | Upregulation | miR-29a | Inhibits VSMC proliferation, migration,and phenotypic switching | Inhibits AD development | Therapeutic target | [38] |
| piRNA-823 | Upregulation | HDAC1 | Promotes VSMC proliferation, migration and phenotypic switching | Promotes AD development | Therapeutic target | [39] |
miRNAs regulating VSMC function
Multiple miRNAs participate in the development of AD by regulating VSMC function through targeting different genes. miR-21 inhibits VSMC apoptosis and promotes proliferation by targeting Phosphatase and Tensin homolog deleted on chromosome ten (PTEN), and simultaneously suppresses VSMC proliferation and promotes vascular remodeling by regulating its target gene SV2C[21]. miR-22 inhibits VSMC apoptosis by targeting p38 during vascular remodeling in AD; its knockdown significantly accelerates disease progression in a mouse model of AD, suggesting the potential therapeutic target value of the MAPK signaling pathway[20]. Wang et al. found that miR-107-5p is highly expressed in acute AD tissues. Overexpression of this miRNA promotes the proliferation and inhibits the apoptosis of rat aortic smooth muscle cells (RASMCs) by targeting ITM2C[22].
In addition, miR-124 is significantly downregulated in the aortic adventitia of clinical AD specimens, and regulates VSMC phenotypic switching by targeting Sp1[23]. miR-134-5p is downregulated in thoracic aortic dissection (TAD) tissues, and inhibits PDGF-BB-induced phenotypic switching and migration of VSMCs, while promoting the expression of differentiation and contractile markers[24]. miR-145 induces the proliferation, migration, and apoptosis of VSMCs by targeting SMAD3, a core mediator of the TGF-β signaling pathway[19]. Furthermore, miR-200c-3p promotes the synthetic phenotypic switching of human aortic smooth muscle cells (HASMCs) by downregulating CASZ1 and inhibiting the RhoA/Egfl7 pathway[25].
lncRNAs regulating VSMC function
lncRNAs affect VSMC function mainly through the ceRNA mechanism or by regulating specific signaling pathways. The lncRNA H19 is aberrantly upregulated in the aorta of AD patients, and exerts its effects by competitively binding to and inhibiting miR-193b-3p. Knockout of H19 effectively inhibits PDGF-BB-induced phenotypic dedifferentiation and migration of HASMCs[26]. SENCR, a myocardin-related pro-differentiation factor for VSMCs, is downregulated in aortic tissues of AD patients. It is inhibited by miR-206 via sponge activity, and its overexpression suppresses the proliferation and migration of HASMCs to maintain the contractile phenotype[27].
Zhang et al. found that HIF1A-AS2 acts as a ceRNA for miR-33b to inhibit the proliferation and migration of VSMCs, and simultaneously promotes phenotypic switching by regulating HMGA2[28]. PVT1 is also involved in disease progression and is upregulated in AD tissues. It reduces the expression of α-SMA and SM22α by targeting miR-27b-3p, thereby promoting the phenotypic switching of VSMCs[29]. Animal studies have shown that overexpression of CDKN2B-AS1 promotes VSMC apoptosis and inhibits their proliferation by acting as a molecular sponge for miR-320d to positively regulate STAT3 expression[30].
The lncRNA PTENP1 is a pseudogene of phosphatase and tensin homolog. Lai et al. found that PTENP1 promotes PTEN expression by acting as a molecular sponge for miR-21, thereby inhibiting downstream Akt signaling and affecting VSMC proliferation and apoptosis[31]. X-Inactive Specific Transcript (XIST) regulates the proliferation and apoptosis of smooth muscle cells by inhibiting miR-17 and its downstream PTEN gene[32]. LINC01278 downregulates ACTG2 by acting as a molecular sponge for miR-500b-5p, promoting smooth muscle phenotypic switching[33]. lncRNA-RP11-465L10.10 is highly expressed in AD tissues, and promotes VSMC phenotypic switching, MMP9 expression, and proliferation/migration via the NF-κB signaling pathway[34].
circRNAs regulating VSMC function
circRNAs, as a class of ncRNAs with stable structure and no free ends, mainly achieve precise regulation of VSMC function by forming a specific regulatory axis or binding functional proteins. The main functional mode of circRNAs is to sponge miRNAs through the ceRNA mechanism, thus forming a complete regulatory axis.
Liu et al. found that in the diseased tissues of AD patients, the expression levels of circ_0000006 and circ_0000160 were significantly upregulated, while the expression of miRNA hsa-let-7e-5p was downregulated. The three together formed a circRNA-miRNA-mRNA regulation axis targeting UBQLN4 gene, which affected the abnormal phenotypic switching, migration and proliferation of VSMC by regulating the expression of UBQLN4, and promoted the pathological progression of aorta from aortic aneurysm (AA) to AD[35]. Other studies have revealed that circ_0022920 is significantly downregulated in AD lesion tissues, and it exerts a protective effect by targeting miR-650. miR-650 negatively regulates TGFβR1 expression via targeting IRF1, and the circ_0022920/miR-650/IRF1/TGFβR1 axis ultimately inhibits the abnormal proliferation and migration of HASMCs, thus delaying AD pathological progression[36]. Cao et al. further clarified its mechanism of action through RNA immunoprecipitation (RIP) experiments, and confirmed that circNRIP1 can enhance the stability of CXCL5 mRNA by competitively binding with insulin-like growth factor 2 binding protein 1 (IGF2BP1), thereby promoting the disease progression of TAD[37]. Xu et al. found that circ-TGFBR2 can act as a molecular sponge of miR-29a, and its inhibitory effect on KLF4 gene can be relieved by adsorbing miR-29a, while the overexpression of circ-TGFBR2 can significantly upregulate the expression of VSMC contractile phenotype markers, downregulate the expression of synthetic phenotype markers, effectively inhibit the abnormal activation of VSMC, thus delaying the pathological progression of AD[38].
Other ncRNAs regulating VSMC function
In addition, researchers have identified a novel small ncRNA, piRNA-823, which is highly expressed in patients with AD. This piRNA directly binds to and inhibits the expression of histone deacetylase 1 (HDAC1), and regulates the acetylation of histone H3 through H3K9ac and H3K27ac modifications, thereby promoting the proliferation, migration, and phenotypic switching of VSMCs[39].
Extracellular matrix dysregulation and associated ncRNA regulation
The ECM is a key component of the aortic wall, mainly composed of collagens type I and III, elastin, proteoglycans, glycosaminoglycans, matrix metalloproteinases (MMPs), and tissue inhibitors of metalloproteinases (TIMPs). These components collectively maintain the elasticity and mechanical stability of the vascular wall[40,41]. ncRNAs mainly disrupt the balance of ECM degradation/synthesis by promoting the overexpression of MMPs (such as MMP-2 and MMP-9) and inhibiting the activity of TIMPs (such as TIMP-1 and TIMP-2). This leads to extensive degradation of collagen and elastin, destroying the structural integrity of the vascular wall, and ultimately promoting the occurrence and development of AD. The roles of ncRNAs in ECM regulation are summarized in Table 2.
ncRNAs influencing AD development by regulating ECM functions
| ncRNA type | Expression status | Target/pathway | Primary regulatory process | Role in AD | Potential application directions | References |
| miR-20a | Upregulation | TIMP-2, MMP-2 | Promotes ECM degradation | Promotes AD development | Therapeutic target | [42] |
| miR-183 | Upregulation | - | Promotes ECM remodeling | Promotes AD development | Therapeutic target | [43] |
| LncRNA-ENSG00000269936 | Upregulation | MAP2K6 | Promotes collagen degradation | Promotes AD development | Therapeutic target | [44] |
| Lnc-C2orf63-4-1 | Downregulation | STAT3 | Inhibits ECM degradation and VSMC apoptosis | Inhibits AD formation | Therapeutic target | [45] |
| hsa_circRNA_101238 | Upregulation | hsa-miR-320a | Promotes ECM degradation | Promotes AD development | Diagnostic/therapeutic target | [46] |
| hsa_circRNA_082317 | Upregulation | ITGA5 | Disrupts ECM-cell adhesion stability | Promotes AD development | Therapeutic target | [47] |
ECM-related miRNAs
Sun et al. confirmed that Nanog homeobox (NANOG) promotes the transcription of miR-20a by binding to the promoter region within the miR-17-92 gene cluster. Upregulated miR-20a expression inhibits TIMP-2 expression, thereby activating MMP-2 and inducing ECM degradation to promote the development of thoracic aortic aneurysm and dissection (AAD)[42]. Wei et al. verified via quantitative real-time polymerase chain reaction (qRT-PCR) and Western blot that miR-183 may participate in ECM remodeling and promote AAD progression by regulating MMP-9 expression in smooth muscle cells[43].
ECM-related lncRNAs
MAP2K6 is a member of the MAPK signaling pathway. Using high-throughput sequencing (HTS), Sun et al. identified the upregulated lncRNA ENSG00000269936, which may target the cis-acting protein-coding gene MAP2K6 to promote ECM degradation[44]. Through HTS analysis and bioinformatics prediction, Zhang et al. identified STAT3 as a downstream target of Lnc-C2orf63-4-1. Downregulation of Lnc-C2orf63-4-1 increases STAT3 expression, promotes Ang II-induced VSMC apoptosis via the STAT3 pathway, and simultaneously significantly reduces Ang II-induced upregulation of MMP-2 and MMP-9. The increased STAT3 expression induced by low Lnc-C2orf63-4-1 expression enhances Ang II-induced VSMC apoptosis, thereby promoting the development of AD[45].
ECM-associated circRNAs
By comparing arterial segments from human patients with type A TAD and healthy donors, researchers found that hsa-miR-320a was downregulated and MMP9 was upregulated in the TAD group, suggesting that hsa_circRNA_101238 may increase MMP9 expression by inhibiting hsa-miR-320a[46]. Liu et al. verified that hsa-circRNA_082317 and ITGA5 were significantly upregulated in AD, while hsa-miR-149-3p was downregulated. hsa-circRNA-082317 may regulate the pathogenesis and progression of AD by competing with hsa-miR-149-3p to mediate ITGA5, a target related to the ECM-receptor interaction pathway and focal adhesion pathway[47].
Inflammatory response and associated ncRNA regulation
AD pathogenesis is closely related to chronic vascular wall inflammation and acute inflammatory storms. Inflammatory cells such as neutrophils and macrophages infiltrate the aortic media, and the expression levels of most MMP genes are significantly elevated in macrophages[48]. These inflammatory cells release a large number of pro-inflammatory cytokines; for example, Interleukin-1β (IL-1β) significantly upregulates the expression of MMP-2 and MMP-9, leading to elastic fiber rupture and ECM degradation[49]. Meanwhile, they induce VSMC dysfunction, ultimately destroying the structural stability of the vascular wall and promoting the formation and progression of dissection. The roles of ncRNAs in inflammatory regulation are summarized in Table 3.
ncRNAs influencing AD onset by regulating inflammatory functions
| ncRNA type | Expression status | Target/pathway | Primary regulatory process | Role in AD | Potential application directions | References |
| miR-181b | Downregulation | Serotonin receptor 6, NF-κB | Inhibits vascular inflammation | Inhibits AD formation | Therapeutic target | [21] |
| miR-30a | Upregulation | MAPK | Promotes vascular inflammation | Promotes AD development | Therapeutic target | [50] |
| miR-127-3p | Upregulation | NF-κB | Promotes vascular inflammation | Promotes AD development | Therapeutic target | [51] |
| lnc-OIP5-AS1 | Upregulation | miR-143-3p | Promotes vascular inflammation and VSMC apoptosis | Promotes AD development | Diagnostic/therapeutic target | [52] |
| hsa_circ_0079929 | Upregulation | hsa_mir_944, NF-κB | Promotes vascular inflammation | Promotes AD onset | Diagnostic/therapeutic target | [53] |
| circNOX4, circAMN1, circUSP3, circPTGR1 | Upregulation | - | Promotes inflammatory infiltration and ECM degradation | Promotes AD development | Diagnostic/therapeutic target | [54] |
Inflammation-associated miRNAs
Using a rat model of AD, Wu et al. found that miR-30a expression was increased in the model group. Inhibition of miR-30a ameliorated the inflammatory response, suggesting that it may participate in inflammation-related AD formation by regulating the MAPK signaling pathway[50]. Using Agilent human miRNA microarray technology, researchers detected that the downstream target gene of miR-181b, 5-hydroxytryptamine receptor 6, can directly inhibit the cyclic adenosine monophosphate (cAMP) signaling pathway. By regulating the expression of protein kinase A (PKA), this mechanism modulates the NF-κB signaling pathway and promotes the release of inflammatory mediators such as IL-8 and IL-1β, thereby participating in the pathogenesis of AD[21].The expression of miR-127-3p is significantly upregulated under vascular injury conditions. Daniel et al. found that miR-127-3p may exert its effects by regulating inflammatory signaling pathways such as NF-κB. Overexpression of miR-127-3p may exacerbate inflammatory infiltration at the site of vascular injury, thereby promoting vascular remodeling[51].
Inflammation-related lncRNAs
Wang found that Lnc-OIP5-AS1 was highly expressed, while miR-143-3p was lowly expressed in aortic tissues of AD patients. Subsequent studies showed that Lnc-OIP5-AS1 indirectly upregulates Tubby bipartite transcription factor (TUB) expression by competitively binding to miR-143-3p, thereby inhibiting the proliferation and promoting the apoptosis of human aortic endothelial cells (HAECs) and HASMCs, suppressing HASMC migration, disrupting the balance of the MMP-2/9 and TIMP-1/2 system, and enhancing the secretion of inflammatory cytokines in human aortic adventitial fibroblasts (HAAFs), ultimately promoting the occurrence and development of AD[52].
Inflammation-related circRNAs
Researchers found that hsa_circ_0079929 is highly expressed and localized in the cytoplasm of human umbilical vein endothelial cells (HUVECs). Knockdown of hsa_circ_0079929 inhibited cell proliferation, invasion, and inflammatory responses. It forms a regulatory pathway together with hsa_miR_944 and PDE4D. Inhibition of hsa_miR_944 activates the NF-κB pathway, indicating that the hsa_circ_0079929/hsa_miR_944/PDE4D pathway may participate in early endothelial injury in AD via NF-κB activation[53].
Liang et al. identified four circRNAs - circNOX4, circAMN1, circUSP3, and circPTGR1 - that are highly expressed in Thoracic Aortic Aneurysm and Dissection (TAAD) tissues and serum. Among them, circNOX4, circAMN1, and circUSP3 are closely related to inflammatory cell infiltration and matrix component loss, while circPTGR1 shows a negative correlation with stromal cell components. These circRNAs promote inflammatory cell infiltration and ECM degradation by interacting with miRNAs, accelerating the progression of TAAD[54].
CLINICAL DIAGNOSIS AND APPLICATIONS
ncRNAs as Biomarkers for early diagnosis of AD
The early diagnosis of acute AD remains challenging. Conventional imaging examinations are costly and often fail to detect early or atypical cases[55]. Accumulating evidence indicates that circulating ncRNAs in the blood exhibit remarkable stability, tissue specificity, and accessibility, making them potential biomarkers for the diagnosis of early and atypical AD[56].
Among these, miRNAs have been the most extensively investigated. Multiple miRNAs that regulate VSMC transformation and inflammatory signaling show significant differential expression in the blood of AD patients. Numerous studies have confirmed that the levels of miRNAs related to VSMC contraction (such as miR-143, miR-145, and members of the miR-29 family) are decreased in the blood of AD patients[57,58], while the levels of inflammation-related miRNAs (including miR-155, miR-221, and miR-222) are elevated[59,60].
For example, the plasma levels of miR-143/145 in patients with acute Stanford type A AD are significantly lower than those in the control group, with reported area under the receiver operating characteristic curve (AUC) values of 0.78-0.89, indicating moderate-to-high diagnostic efficiency. Similarly, elevated circulating levels of miR-155 and miR-221/222 are correlated with inflammatory activity and disease severity, suggesting their combined diagnostic and pathophysiological significance[59,61,62].
In addition to miRNAs, lncRNAs and circRNAs also show differential expression. For instance, lncRNAs related to ECM remodeling (such as MALAT1) and circRNAs associated with vascular integrity (including circRNA_0001806 and circRNA_002178) are aberrantly expressed in AD patients[54,63]. Taken together, these findings indicate that ncRNAs have important potential as biomarkers for the early diagnosis of AD.
Clinical therapeutic targets
ncRNAs affect the development of AD by regulating the phenotypic switching of VSMCs, ECM degradation and inflammatory activation. Therefore, targeting regulatory related ncRNAs can delay, inhibit or even repair the pathological state of aortic fragility, thereby maintaining vascular stability. For example, supplementing miR-143/145 gene cluster can stabilize the contractile phenotype of VSMCs and reduce aortic wall degeneration[54,57], while inhibiting miR-323a-3p and miR-449b-5p can alleviate inflammatory injury and abnormal VSMC proliferation[64]. The expression and function of lncRNAs and circRNAs can be modulated using nucleic acid-based strategies including antisense oligonucleotides (ASOs), small interfering RNA (siRNA), and CRISPR-Cas9 gene editing technology. For instance, targeting the lncRNA MALAT1 reduces ECM degradation, while restoration of lncRNA GAS5 expression alleviates vascular inflammatory stress and VSMC apoptosis[65,66].
Research based on multi-omics integration and RNA association networks has revealed that the ceRNA regulatory network formed by ncRNAs can maintain vascular microenvironment homeostasis through multi-dimensional regulation, which has become the core basis for molecular typing and target screening of AD. Meanwhile, CRISPR-Cas9 gene editing technology enables precise knockout or overexpression of ncRNA targets, providing an efficient genetic manipulation tool for the modulation of the above core regulatory network[67]. In addition, the restoration of protective circRNA expression can enhance aortic intimal stability[68].
Notably, deep learning algorithms can construct highly accurate models for AD molecular subtyping and therapeutic efficacy prediction, based on the sequence characteristics, target correlation, and expression profile data of ncRNAs. These models can provide intelligent analytical tools for the screening of CRISPR-Cas9 editing targets, as well as the personalized optimization of ncRNA-targeted therapies[69].
These studies are still in the preclinical stage, but the cross-integration of multi-omics analysis, RNA association research, gene editing technology and deep learning is expected to markedly improve the precision and efficacy of ncRNA-targeted therapies. This integrated approach will also provide novel research directions and technical support for slowing disease progression in patients with early-stage AD, and for the development of new targeted treatments for Stanford type B AD.
ncRNAs as prognostic biomarkers for AD
Alterations in circulating ncRNAs not only reflect disease onset but also correlate with postoperative complications and long-term survival rates. Elevated levels of MALAT1 and miR-155 are closely associated with an increased risk of re-dissection, adverse events, and reduced survival rates[63,70], indicating that these molecules reflect persistent molecular instability of the aortic wall. Conversely, elevated levels of protective ncRNAs such as miR-143/145 and GAS5 predict favorable vascular remodeling and a reduced recurrence rate[54,65].
Taken together, these findings suggest that ncRNAs constitute a multidimensional biomarker system that can provide a basis for the diagnosis, treatment decision-making, and prognostic assessment of AD. The integration of ncRNAs into clinical workflows is expected to enable earlier disease detection, more accurate risk stratification, and more effective personalized treatment regimens.
Current detection technologies for ncRNA biomarkers
The translation of ncRNA biomarkers into clinical diagnostic tools is highly dependent on reliable detection platforms. The current main technologies include:
qRT-PCR: The most widely used clinical validation tool due to its high sensitivity and specificity. However, standardization strategies remain inconsistent, and pre-analytical variability (sample processing, hemolysis) significantly affects reproducibility[59].
Digital PCR (dPCR): Provides high-precision absolute quantitative analysis and is being widely applied to the detection of low-abundance circulating ncRNAs. It has broad prospects for clinical scalability, but is limited by cost and equipment availability[71,72].
Next-generation sequencing (NGS): Enables comprehensive profiling of biomarkers. Despite its powerful capabilities, high costs, complex bioinformatics processing, and the lack of standardized workflows currently limit its routine clinical application[16].
Microarray platforms: Suitable for large-scale screening, but have low sensitivity for lowly expressed ncRNAs and are gradually being replaced by sequencing technologies[59].
Challenges in clinical translation
Although these biomarkers show considerable translational potential, their clinical application still faces multiple key barriers, including the lack of large-scale multi-center validation cohorts, insufficient control of pre-analytical variability and standardized workflows, significant biological heterogeneity between disease subtypes, and regulatory and compliance limitations of nucleic acid diagnostics. Meanwhile, their cost-effectiveness relative to existing imaging diagnostic pathways remains uncertain.
Effective advancement of clinical translation urgently requires expanding the evidence base through multi-center collaborative clinical studies, establishing a unified and reproducible standardized operating system, and integrating molecular biomarkers into a multimodal diagnostic framework to improve the reliability and clinical feasibility of overall diagnosis.
SUMMARY AND OUTLOOK
Current research and emerging directions
Current research on ncRNAs in AD has made phased progress. Research hotspots focus on three major molecular categories: miRNAs, lncRNAs, and circRNAs. These molecules have been confirmed to participate in the pathogenesis of AD by regulating VSMC phenotypic switching, ECM homeostasis, and vascular inflammatory responses. For example, miR-20a accelerates ECM degradation by inhibiting TIMP-2 and promoting MMP-2 activation[42], the lncRNA SENCR regulates VSMC proliferation and apoptosis by targeting miR-206[27].
Meanwhile, ncRNAs with tissue-specific differential expression have shown potential as diagnostic biomarkers for AD, such as hsa-miRNA-4787-5p[73] and miR-15a[74], opening up new avenues for the early detection of AD. In addition, exploring the role of ncRNAs in the regulation of metabolic and signaling pathways has become a key breakthrough point for deciphering the pathophysiological mechanisms of AD.
Technical limitations and gaps in clinical validation
Current research on ncRNAs in AD is still limited by key technical bottlenecks, including insufficient cellular resolution, missing spatial expression information, and low in vivo delivery efficiency. Traditional bulk transcriptome analysis fails to effectively reveal the cellular heterogeneity of the aortic wall and the cell-specific regulatory networks of ncRNAs.
Future research should establish a closed-loop research framework of “single-cell resolution-spatial localization-functional validation”: (1) identifying key ncRNA regulatory modules via single-cell multi-omics; (2) clarifying their hierarchical distribution in the aortic wall microenvironment in combination with spatial transcriptomics; (3) verifying their causal roles in VSMC phenotypic switching and inflammatory remodeling using targeted intervention technologies. Meanwhile, cross-modal computational models should be introduced to integrate high-dimensional data and screen regulatory nodes with translational potential[75].
At the therapeutic level, two core breakthrough directions should be prioritized. First, we need to break through the delivery limitations of nucleic acid drugs in the high-shear force vascular environment, and develop cell-targeted, biomimetic, and microenvironment-responsive carrier systems. Second, the stability of biomarkers and the standardization of detection methods should be verified through multi-center clinical cohorts, so as to promote the translation of ncRNA research from mechanistic correlation to precise clinical intervention.
Future research priorities and directions
Constructing Precise Expression Maps
Future efforts should integrate single-cell sequencing, spatial transcriptomics, and artificial intelligence (AI) to systematically map the dynamic expression profiles of ncRNAs across different AD subtypes and disease stages. This will clarify the cellular localization and expression characteristics of key ncRNAs, enabling the selection of highly specific and sensitive biomarkers to support early diagnosis and risk stratification in AD.
In-depth exploration of regulatory mechanisms
Research should focus on the interaction networks between ncRNAs and DNA, RNA, and proteins to decipher their molecular mechanisms in core pathological processes such as VSMC phenotypic switching and metabolic reprogramming. This includes elucidating the regulatory pathways of lncRNA-encoded functional peptides.
Developing efficient targeted therapeutics
Based on mechanistically validated targets, researchers should advance ncRNA-targeted drug development by designing nucleic acid therapeutics or small-molecule modulators. Prioritize addressing drug delivery efficiency, tissue specificity, and safety concerns. Achieve precise ncRNA enrichment at diseased aortic sites through optimized delivery systems. Concurrently conduct large-scale clinical studies to validate therapeutic efficacy, ultimately bridging the gap from mechanistic research to clinical translation.
Ethical and safety considerations in the clinical application of ncRNAs
With the extension of ncRNAs into the field of clinical diagnosis and treatment, their ethical and safety challenges have become increasingly prominent. ncRNAs have multi-pathway regulatory properties, and their intervention may cause off-target effects or long-term transcriptional remodeling. Therefore, it is necessary to systematically evaluate multi-organ toxicity and long-term safety in preclinical studies.
Nucleic acid delivery materials may also induce immune responses, especially in vascular diseases with an inflammatory background, and the risk should be reduced through carrier optimization and dose control. At the same time, ncRNA diagnosis and treatment rely on the collection of high-dimensional molecular data, involving ethical issues such as privacy protection, data sharing, and informed consent, which urgently require the establishment of a standardized governance framework.
At the clinical implementation level, the principles of evidence-based and phased validation should be followed, and multidisciplinary ethical review and risk monitoring should be strengthened. Synchronous integration of safety assessment, ethical norms, and technology development into the research design is a fundamental prerequisite for ensuring the sustainable advancement of the translational application of ncRNAs.
DECLARATIONS
Authors’ contributions
Conceptualization, manuscript drafting, and manuscript polishing: Yang J
Data collection, manuscript drafting, and manuscript revision: Wu W
Assisted in literature collection and data sorting: Duan Y, Pan M, Wu L, Wang S, Huang J, Du J
Supervision, project administration, and final review of the manuscript: Liu Q
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported by the National Natural Science Foundation of China (Grant No. 82271923) and the National Natural Science Foundation of China (Grant No. 8224100738). The funders had no role in the design of the study, data collection and analysis, interpretation of the results, writing of the manuscript, or the decision to publish.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Isselbacher EM, Preventza O, Hamilton Black J 3rd, et al.; Peer Review Committee Members. 2022 ACC/AHA guideline for the diagnosis and management of aortic disease: a report of the American Heart Association/American College of Cardiology Joint Committee on Clinical Practice Guidelines. Circulation. 2022;146:e334-482.
2. Erbel R, Aboyans V, Boileau C, et al. ; Grupa Robocza Europejskiego Towarzystwa Kardiologicznego (ESC) do spraw rozpoznawania i leczenia chorób aorty. 2014 ESC Guidelines on the diagnosis and treatment of aortic diseases. Kardiol Pol. 2014;72:1169-252.
3. Rensen SS, Doevendans PA, van Eys GJ. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Neth Heart J. 2007;15:100-8.
4. Ikonomidis JS, Jones JA, Barbour JR, et al. Expression of matrix metalloproteinases and endogenous inhibitors within ascending aortic aneurysms of patients with Marfan syndrome. Circulation. 2006;114:I365-70.
5. He R, Guo DC, Estrera AL, et al. Characterization of the inflammatory and apoptotic cells in the aortas of patients with ascending thoracic aortic aneurysms and dissections. J Thorac Cardiovasc Surg. 2006;131:671-8.
6. The Writing Committee of the Report on Cardiovascular Health and Diseases in China. Report on cardiovascular health and diseases in China 2021: an updated summary. Biomed Environ Sci. 2022;35:573-603.
7. Fattori R, Cao P, De Rango P, et al. Interdisciplinary expert consensus document on management of type B aortic dissection. J Am Coll Cardiol. 2013;61:1661-78.
8. Hagan PG, Nienaber CA, Isselbacher EM, et al. The international registry of acute aortic dissection (IRAD): new insights into an old disease. JAMA. 2000;283:897-903.
9. Estrera AL, Miller CC 3rd, Safi HJ, et al. Outcomes of medical management of acute type B aortic dissection. Circulation. 2006;114:I384-9.
10. Evangelista A, Isselbacher EM, Bossone E, et al. ; IRAD Investigators. Insights from the international registry of acute aortic dissection: a 20-year experience of collaborative clinical research. Circulation. 2018;137:1846-60.
14. Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495:333-8.
15. Elia L, Quintavalle M, Zhang J, et al. The knockout of miR-143 and -145 alters smooth muscle cell maintenance and vascular homeostasis in mice: correlates with human disease. Cell Death Differ. 2009;16:1590-8.
16. Ding W, Liu Y, Su Z, Li Q, Wang J, Gao Y. Emerging role of non-coding RNAs in aortic dissection. Biomolecules. 2022;12:1336.
17. Wang XJ, Huang B, Zhang L, et al. Differential gene expression in aortic tissues between patients with acute aortic dissection and normal controls. Chin J Mol Cardiol. 2014;14. 812-5. (in Chinese).
18. Guo DC, Regalado ES, Minn C, et al. Familial thoracic aortic aneurysms and dissections: identification of a novel locus for stable aneurysms with a low risk for progression to aortic dissection. Circ Cardiovasc Genet. 2011;4:36-42.
19. Huang W, Huang C, Ding H, et al. Involvement of miR-145 in the development of aortic dissection via inducing proliferation, migration, and apoptosis of vascular smooth muscle cells. J Clin Lab Anal. 2020;34:e23028.
20. Xiao Y, Sun Y, Ma X, et al. MicroRNA-22 inhibits the apoptosis of vascular smooth muscle cell by targeting p38MAPKα in vascular remodeling of aortic dissection. Mol Ther Nucleic Acids. 2020;22:1051-62.
21. Zou S. Analysis of microRNA expression and regulatory mechanisms in Stanford type A aortic dissection. Dissertation, University of South China, Hunan, 2015. (in Chinese). Available from: https://med.wanfangdata.com.cn/Paper/Detail?dbid=WF_XW&id=DegreePaper_D774884. [Last accessed on 2 June 2026].
22. Wang Z, Zhuang X, Chen B, Feng D, Li G, Wei M. The role of miR-107 as a potential biomarker and cellular factor for acute aortic dissection. DNA Cell Biol. 2020;39:1895-906.
23. Tang Y, Yu S, Liu Y, Zhang J, Han L, Xu Z. MicroRNA-124 controls human vascular smooth muscle cell phenotypic switch via Sp1. Am J Physiol Heart Circ Physiol. 2017;313:H641-9.
24. Wang Y, Dong CQ, Peng GY, et al. MicroRNA-134-5p regulates media degeneration through inhibiting VSMC phenotypic switch and migration in thoracic aortic dissection. Mol Ther Nucleic Acids. 2019;16:284-94.
25. Shao Y. The role of miR200c-3p in regulating endothelial cell apoptosis and proliferation and vascular smooth muscle cell phenotypic switching in aortic dissection. Dissertation, Chongqing Medical University, Chongqing, 2022. (in Chinese).
26. Ren M, Wang T, Wei X, et al. LncRNA H19 regulates smooth muscle cell functions and participates in the development of aortic dissection through sponging miR-193b-3p. Biosci Rep. 2021;41.
27. Song Y, Wang T, Mu C, Gui W, Deng Y, Ma R. LncRNA SENCR overexpression attenuated the proliferation, migration and phenotypic switching of vascular smooth muscle cells in aortic dissection via the miR-206/myocardin axis. Nutr Metab Cardiovasc Dis. 2022;32:1560-70.
28. Zhang K, Qi Y, Wang M, Chen Q. Long non-coding RNA HIF1A-AS2 modulates the proliferation, migration, and phenotypic switch of aortic smooth muscle cells in aortic dissection via sponging microRNA-33b. Bioengineered. 2022;13:6383-95.
29. Li S, Zhao X, Cheng S, Li J, Bai X, Meng X. Downregulating long non-coding RNA PVT1 expression inhibited the viability, migration and phenotypic switch of PDGF-BB-treated human aortic smooth muscle cells via targeting miR-27b-3p. Hum Cell. 2021;34:335-48.
30. Zhao X, Cheng S, Li S, Li J, Bai X, Xi J. CDKN2B-AS1 aggravates the pathogenesis of human thoracic aortic dissection by sponge to miR-320d. J Cardiovasc Pharmacol. 2020;76:592-601.
31. Lai Y, Li J, Zhong L, et al. The pseudogene PTENP1 regulates smooth muscle cells as a competing endogenous RNA. Clin Sci. 2019;133:1439-55.
32. Zhang X, Wu H, Mai C, Qi Y. Long noncoding RNA XIST/miR-17/PTEN axis modulates the proliferation and apoptosis of vascular smooth muscle cells to affect stanford type a aortic dissection. J Cardiovasc Pharmacol. 2020;76:53-62.
33. Wang W, Liu Q, Wang Y, et al. LINC01278 sponges miR-500b-5p to regulate the expression of ACTG2 to control phenotypic switching in human vascular smooth muscle cells during aortic dissection. J Am Heart Assoc. 2021;10:e018062.
34. Lin Y, Huang H, Yu Y, et al. Long non-coding RNA RP11-465L10.10 promotes vascular smooth muscle cells phenotype switching and MMP9 expression via the NF-κB pathway. Ann Transl Med. 2021;9:1776.
35. Liu Y, Wang L, Lei D, et al. Circ_0000006 and circ_0000160 regulate hsa-let-7e-5p/UBQLN4 axis in aortic dissection progression. PLoS One. 2024;19:e0304668.
36. Huang MX, Piao HL, Wang Y, et al. Circ_0022920 maintains the contractile phenotype of human aortic vascular smooth muscle cells via sponging microRNA-650 and promoting transforming growth factor beta receptor 1 expression in angiotensin II-induced models for aortic dissection. J Am Heart Assoc. 2023;12:e027425.
37. Cao X, Fang H, Zhou L. CircNRIP1 promotes proliferation, migration and phenotypic switch of Ang II-induced HA-VSMCs by increasing CXCL5 mRNA stability via recruiting IGF2BP1. Autoimmunity. 2024;57:2304820.
38. Xu Z, Zhong K, Guo G, et al. circ_TGFBR2 inhibits vascular smooth muscle cells phenotypic switch and suppresses aortic dissection progression by sponging miR-29a. J Inflamm Res. 2021;14:5877-90.
39. Li M, Li G, Yang Y, et al. piRNA-823 is a novel potential therapeutic target in aortic dissection. Pharmacol Res. 2023;196:106932.
40. Xu J, Shi GP. Vascular wall extracellular matrix proteins and vascular diseases. Biochim Biophys Acta. 2014;1842:2106-19.
41. Tian T, Luo F, Zhao LP, et al. Research progress on the role and mechanism of extracellular matrix in aortic aneurysm and aortic dissection. Chin J Clin Thorac Cardiovasc Surg. 2024;31. 1376-84. (in Chinese).
42. Sun YY. The role and mechanism of NANOG transcriptional regulation of miR-20a in promoting extracellular matrix degradation in the pathogenesis of thoracic aortic dissection. Dissertation, Naval Medical University, Shanghai, 2022.(in Chinese).
43. Wei Y, Yuan M, Wei M, et al. Study on the role and mechanism of microRNA-183 in extracellular matrix remodeling of acute aortic dissection. Chin J Mod Med. 2022;32:28-32.
44. Sun J, Chen G, Jing Y, et al. LncRNA expression profile of human thoracic aortic dissection by high-throughput sequencing. Cell Physiol Biochem. 2018;46:1027-41.
45. Zhang S, Zhao S, Han X, et al. Lnc-C2orf63-4-1 confers VSMC homeostasis and prevents aortic dissection formation via STAT3 interaction. Front Cell Dev Biol. 2021;9:792051.
46. Zou M, Huang C, Li X, et al. Circular RNA expression profile and potential function of hsa_circRNA_101238 in human thoracic aortic dissection. Oncotarget. 2017;8:81825-37.
47. Liu DB, He YF, Chen GJ, et al. Construction of a circRNA-Mediated ceRNA network reveals novel biomarkers for aortic dissection. Int J Gen Med. 2022;15:3951-64.
48. Liu X, Chen W, Zhu G, et al. Single-cell RNA sequencing identifies an Il1rn+/Trem1+ macrophage subpopulation as a cellular target for mitigating the progression of thoracic aortic aneurysm and dissection. Cell Discov. 2022;8:11.
49. Guo LL, Wu MT, Zhang LW, et al. Blocking interleukin-1 beta reduces the evolution of thoracic aortic dissection in a rodent model. Eur J Vasc Endovasc Surg. 2020;60:916-24.
50. Wu YW, Hu B, Guo XD, Fu Q, Zou ZJ. Effect of microRNA-30a regulation of mitogen-activated protein kinase pathway on aortic coarctation in rats. Acta Anat Sin 2024;55:222-8. (in Chinese).
51. Daniel F, Kalies K, Hehl L, et al. MicroRNA-127-3p regulates inflammatory response in injury-induced vascular remodeling and cellular senescence in vascular smooth muscle cells through mTor. Cardiovasc Res. 2024;120:cvae088.006.
52. Wang P. The mechanism of Lnc-OIP5-AS1 competitively binding to miR-143-3p in the pathogenesis of aortic dissection. Dissertation, Wuhan University, Wuhan, 2021. (in Chinese).
53. Wang K. Circular RNA hsa_circ_0079929 promotes endothelial inflammation in aortic dissection by activating PDE4D. Dissertation, Qingdao University, Shandong, 2023. (in Chinese).
54. Liang Q, Zhou Z, Li H, et al. Identification of pathological-related and diagnostic potential circular RNAs in Stanford type A aortic dissection. Front Cardiovasc Med. 2022;9:1074835.
55. Pape LA, Awais M, Woznicki EM, et al. Presentation, diagnosis, and outcomes of acute aortic dissection: 17-year trends from the international registry of acute aortic dissection. J Am Coll Cardiol. 2015;66:350-8.
56. Barile L, Vassalli G. Exosomes: therapy delivery tools and biomarkers of diseases. Pharmacol Ther. 2017;174:63-78.
57. Liao M, Zou S, Weng J, et al. A microRNA profile comparison between thoracic aortic dissection and normal thoracic aorta indicates the potential role of microRNAs in contributing to thoracic aortic dissection pathogenesis. J Vasc Surg. 2011;53:1341-1349.e3.
58. Boon RA, Seeger T, Heydt S, et al. MicroRNA-29 in aortic dilation: implications for aneurysm formation. Circ Res. 2011;109:1115-9.
59. Xu Z, Wang Q, Pan J, et al. Characterization of serum miRNAs as molecular biomarkers for acute Stanford type A aortic dissection diagnosis. Sci Rep. 2017;7:13659.
60. Chai T, Yang X, Zhuang X, et al. Upregulation of miR-222-3p alleviates the symptom of aortic dissection through targeting STAT3. Life Sci. 2022;310:121051.
61. Zhang D, Zhao X, Wang B, Liu X, Aizezi A, Ma X. Circulating exosomal miRNAs as novel biomarkers for acute aortic dissection: a diagnostic accuracy study. Medicine. 2023;102:e34474.
62. Chen H, Li Y, Li Z, Shi Y, Zhu H. Diagnostic biomarkers and aortic dissection: a systematic review and meta-analysis. BMC Cardiovasc Disord. 2023;23:497.
63. Michalik KM, You X, Manavski Y, et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth. Circ Res. 2014;114:1389-97.
64. Rodor J, Klimi E, Brown SD, et al. Functional screening identifies miRNAs with a novel function inhibiting vascular smooth muscle cell proliferation. Mol Ther. 2025;33:615-30.
65. Li Y, Maegdefessel L. Non-coding RNA contribution to thoracic and abdominal aortic aneurysm disease development and progression. Front Physiol. 2017;8:429.
66. He X, Wang S, Li M, et al. Long noncoding RNA GAS5 induces abdominal aortic aneurysm formation by promoting smooth muscle apoptosis. Theranostics. 2019;9:5558-76.
67. Tamblin-Hopper P, Kiss-Toth E, Sudbery I, et al. The potential therapeutic applications of long non-coding RNAs. J Transl Genet Genom. 2024;8:225-43.
68. He X, Li X, Han Y, et al. CircRNA Chordc1 protects mice from abdominal aortic aneurysm by contributing to the phenotype and growth of vascular smooth muscle cells. Mol Ther Nucleic Acids. 2022;27:81-98.
69. Dogra S, Kouznetsova VL, Kesari S, Tsigelny IF. Development of a miRNA-based deep learning model for autism spectrum disorder diagnosis. Adv Technol Neurosci. 2025;2:72-6.
70. Zhang RL, Wang WM, Li JQ, et al. The role of miR-155 in cardiovascular diseases: potential diagnostic and therapeutic targets. Int J Cardiol Cardiovasc Risk Prev. 2025;24:200355.
71. D’Alessandra Y, Valerio V, Moschetta D, et al. Extraction-free absolute quantification of circulating miRNAs by chip-based digital PCR. Biomedicines. 2022;10:1354.
72. Ferracin M, Negrini M. Quantification of circulating microRNAs by droplet digital PCR. In: Karlin-neumann G, Bizouarn F, editors. Digital PCR. New York: Springer; 2018. pp. 445-57.
73. Wang L, Zhang S, Xu Z, Zhang J, Li L, Zhao G. The diagnostic value of microRNA-4787-5p and microRNA-4306 in patients with acute aortic dissection. Am J Transl Res. 2017;9:5138-49.
74. Dong J, Bao J, Feng R, et al. Circulating microRNAs: a novel potential biomarker for diagnosing acute aortic dissection. Sci Rep. 2017;7:12784.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0











Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].