




Glycosylated RNAs in extracellular vesicles: a new layer of RNA sorting and intercellular communication
 0
0 
MAIN TEXT
The selective sorting of specific RNAs into exosomes remains a central, unresolved question in extracellular RNA biology. Previous studies have largely emphasized RNA-binding proteins and sequence motifs as determinants of RNA sorting, while emerging evidence suggests that RNA chemical modifications and cellular context may also play important roles in shaping extracellular vesicle (EV) RNA composition[1]. However, whether chemical modifications of RNA itself can act as intrinsic sorting signals has remained unclear. Recent advances in the identification and characterization of glycosylated RNAs (glycoRNAs) have begun to reveal their broader biological relevance beyond their initial discovery[2].
A recent study published in Nature Cell Biology establishes a novel conceptual link between RNA chemistry and intercellular communication via extracellular vesicles, specifically highlighting the role of RNA glycosylation[3]. Using metabolic labeling with the azide-modified sugar analog Ac4GalNAz[4], the authors enriched glycoRNAs from living cells and uncovered a striking distribution pattern: glycoRNAs were not detected at the plasma membrane or in the nucleus, but were instead enriched in membrane-associated cytoplasmic compartments and within the lumen of exosomes. This finding contrasts with earlier reports describing cell surface-associated glycoRNAs[5], suggesting that glycoRNAs may comprise functionally distinct subpopulations defined by their subcellular localization: (1) cell-surface glycoRNAs, which are displayed on the outer leaflet of the plasma membrane and likely mediate direct cell-cell interactions; (2) exosome-luminal glycoRNAs, the focus of this study, which are actively sorted into the vesicle lumen via the Endosomal Sorting Complex Required for Transport (ESCRT) pathway and packaged for protected, long-range delivery; and (3) a putative third class of exosome-surface-associated glycoRNAs that may be transiently or peripherally associated with the exosomal membrane exterior and whose roles remain to be resolved. This spatial partitioning implies a functional divergence where surface-exposed glycoRNAs could play immediate roles in immune recognition or cell adhesion, while luminal glycoRNAs are specialized for intercellular communication.
While the metabolic labeling and click chemistry approach provides a powerful strategy for glycoRNA enrichment, a methodological caveat warrants attention. Glycoprotein co-purification may contribute to glycan signals in glycoRNA preparations. Although RNase and proteinase K treatments were applied to reduce protein contamination, residual glycoproteins or glycan-binding proteins cannot be fully excluded. Future validation using stringent detergent washes, mass spectrometry-based linkage characterization, glycosidase digestion, or orthogonal sugar analog labeling would help confirm direct RNA-glycan conjugation.
Notably, the identified glycoRNAs are predominantly small non-coding RNAs (< 200 nucleotides), including snoRNAs, snRNAs, and Y RNAs. This size bias raises important mechanistic questions. One possibility is that short RNAs are more compatible with vesicle loading machinery or preferentially recognized by RNA-binding adaptors involved in cargo selection. Alternatively, glycosylation itself may be structurally more accessible on compact RNA species, thereby favoring smaller transcripts[1]. In addition, glycoRNAs display resistance to RNase A digestion, suggesting that glycosylation may enhance RNA stability and facilitate their persistence in the extracellular environment.
Mechanistically, disruption of exosome biogenesis, either through genetic inhibition of ESCRT components such as HRS[6,7] or pharmacological blockade using GW4869[8] and manumycin A[9], led to a marked reduction of glycoRNAs in exosomes, accompanied by their intracellular accumulation. These findings support a model in which glycoRNAs are actively sorted into exosomes via canonical vesicle biogenesis pathways.
Intriguingly, the study also points to a functional interplay between RNA and protein glycosylation networks. Perturbation of protein glycosylation pathways, including knockdown of GALE or GNE[10] and inhibition of oligosaccharyltransferase activity[11], significantly reduced glycoRNA levels. This observation suggests that RNA and protein glycosylation may share common metabolic substrates and regulatory pathways. Such coordination raises the possibility that cellular metabolic states globally influence glycosylation-dependent cargo selection, given that nucleotide sugar metabolism and glycosylation pathways are tightly integrated within cellular networks[12].
Beyond mechanistic insight, glycoRNAs are capable of intercellular transfer via exosomes. Exosomes derived from labeled donor cells are efficiently taken up by recipient cells, enabling rapid delivery of glycoRNAs. Moreover, the composition of exosomal glycoRNAs dynamically changes during stem cell differentiation, suggesting that RNA glycosylation and vesicle-mediated sorting are responsive to cellular state and may participate in developmental or adaptive processes.
The existence of glycoRNAs in distinct subcellular compartments further raises important questions regarding their biological functions. Surface-exposed glycoRNAs may participate in immune recognition or cell-cell interactions, whereas exosome-associated glycoRNAs may mediate long-range intercellular communication. Differences in glycan structures, RNA species, and interacting partners may further diversify their functional roles across cellular contexts.
CONCLUSION AND FUTURE PERSPECTIVES
This study identifies RNA glycosylation as a novel molecular feature potentially associated with selective RNA sorting into exosomes, thereby adding a new layer of regulation to extracellular RNA biology. By linking RNA chemical modification to vesicle-mediated communication, these findings expand the current framework of RNA sorting beyond protein-centric models. However, while these findings strongly associate RNA glycosylation with exosomal enrichment, definitive proof that glycosylation per se serves as an intrinsic and sufficient sorting signal awaits future functional studies that directly manipulate specific glycoRNA structures.
Several key questions remain to be addressed in future research. First, the exact chemical linkage between glycans and RNAs, whether N-linked, O-linked, or through other covalent bonds, remains to be precisely elucidated. Second, the enzymatic machinery responsible for catalyzing RNA glycosylation, including the identification of the specific glycosyltransferases and their substrate recognition mechanisms, is largely unknown. Third, beyond the sorting process, the biological functions of glycoRNAs following exosomal transfer to recipient cells are a critical frontier. It is imperative to determine whether and how glycoRNAs influence recipient cell physiology, such as modulating immune responses, altering signaling pathways, or contributing to disease progression. Furthermore, the determinants of substrate specificity, particularly the preference for small non-coding RNAs, require further investigation. Future studies should also explore how metabolic and signaling networks coordinate RNA and protein glycosylation, particularly in the context of glycosylation pathway integration and cellular nutrient states.
From a translational perspective, glycoRNAs may represent a new class of biomarkers or therapeutic targets in diseases involving extracellular vesicles. Overall, these findings open new avenues at the intersection of RNA biology, glycobiology, and intercellular communication [Figure 1].
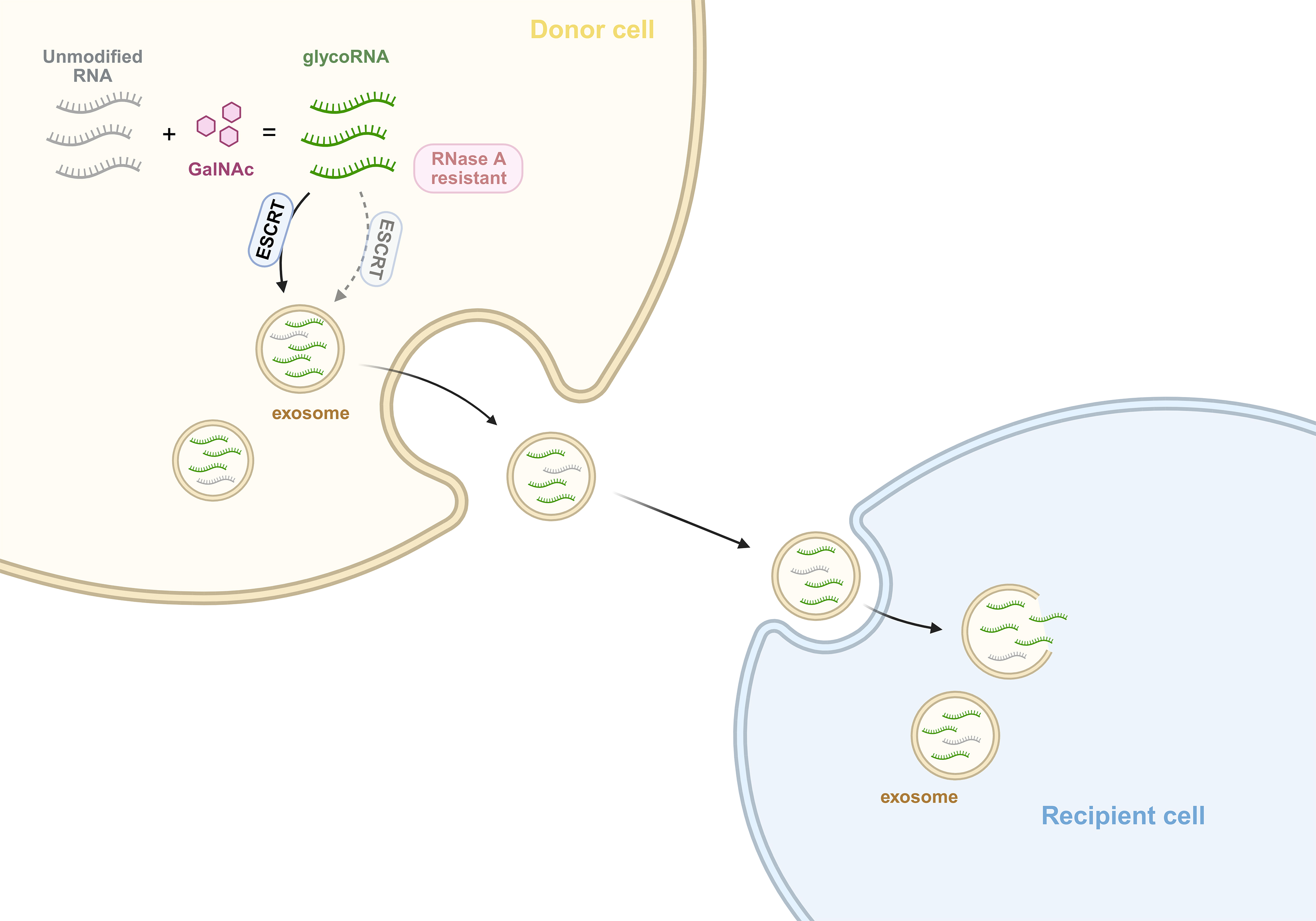
Figure 1. Glycosylation regulates exosomal RNA sorting and intercellular transfer. In donor cells, a subset of small non-coding RNAs undergoes GalNAc-dependent glycosylation, which is associated with their preferential enrichment into exosomes during biogenesis. This specific cargo-loading process depends on canonical exosome biogenesis pathways, including both ESCRT-dependent and ESCRT-independent mechanisms. Subsequently, these glycoRNAs are encapsulated within the exosomal lumen. Exosomes containing glycoRNAs can be released and taken up by recipient cells, facilitating the intercellular transfer of glycoRNAs. Created in BioRender. Wu, Z. (2026) https://BioRender.com/cv1j34x. GalNAc: N-acetylgalactosamine; RNA: ribonucleic acid; ESCRT: endosomal sorting complex required for transport; glycoRNA: glycosylated RNA.
DECLARATIONS
Authors’ contributions
Responsible for the specific writing and drawing: Wu Z
Drafted the outline and revised the manuscript: Cai Z
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool ChatGPT (version 1.2026.051, released 2026-3) was used solely for grammar revision. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
This work was supported by the National Natural Science Foundation of China (82130053 and 82371751).
Conflicts of interest
Cai Z is an Editorial Board Member of the journal Extracellular Vesicles and Circulating Nucleic Acids. Cai Z was not involved in any steps of editorial processing, notably including reviewers’ selection, manuscript handling and decision making. Wu Z declares that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Yadav A, Sharma A, Gavande PV, et al. Mapping GlycoRNAs on an exosomal surface. J Am Chem Soc. 2026;148:2739-48.
2. Kegel NB, Yilmaz Demirel N, Glatter T, Höfer K, Kaufmann A, Bauer S. Proteins are a source of glycans found in preparations of glycoRNA. Exp Mol Med. 2025;57:2505-16.
3. Sharma S, Jiao X, Yang J, Kwan KY, Kiledjian M. Extracellular exosomal RNAs are glyco-modified. Nat Cell Biol. 2025;27:983-91.
4. Boyce M, Carrico IS, Ganguli AS, et al. Metabolic cross-talk allows labeling of O-linked beta-N-acetylglucosamine-modified proteins via the N-acetylgalactosamine salvage pathway. Proc Natl Acad Sci U S A. 2011;108:3141-6.
5. Flynn RA, Pedram K, Malaker SA, et al. Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell. 2021;184:3109-24.e22.
6. Tamai K, Tanaka N, Nakano T, et al. Exosome secretion of dendritic cells is regulated by Hrs, an ESCRT-0 protein. Biochem Biophys Res Commun. 2010;399:384-90.
7. Han QF, Li WJ, Hu KS, et al. Exosome biogenesis: machinery, regulation, and therapeutic implications in cancer. Mol Cancer. 2022;21:207.
8. Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9:654-9.
9. Datta A, Kim H, Lal M, et al. Manumycin A suppresses exosome biogenesis and secretion via targeted inhibition of Ras/Raf/ERK1/2 signaling and hnRNP H1 in castration-resistant prostate cancer cells. Cancer Lett. 2017;408:73-81.
10. Akella NM, Ciraku L, Reginato MJ. Fueling the fire: emerging role of the hexosamine biosynthetic pathway in cancer. BMC Biol. 2019;17:52.
11. Lopez-Sambrooks C, Shrimal S, Khodier C, et al. Oligosaccharyltransferase inhibition induces senescence in RTK-driven tumor cells. Nat Chem Biol. 2016;12:1023-30.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].